 2008, Vol. 44
2008, Vol. 44文章信息
- 徐嘉, 费世民, 何亚平, 蒋俊明, 陈秀明, 郭志华, 涂代伦.
- Xu Jia, Fei Shimin, He Yaping, Jiang Junming, Chen Xiuming, Guo Zhihua, Tu Dailun
- 川西南山地云南松林窗更新特征
- Regeneration Characteristics of Pinus yunnanensis in Gaps on the Mountainous Area in Southwestern Sichuan Province, China
- 林业科学, 2008, 44(9): 7-12.
- Scientia Silvae Sinicae, 2008, 44(9): 7-12.
-
文章历史
- 收稿日期:2007-06-10
-
作者相关文章



2. 四川省林业科学研究院 成都 610081;
3. 中国林业科学研究院森林生态环境与保护研究所 北京 100091;
4. 攀枝花市盐边林业局 盐边 617002
2. Sichuan Academy of Forestry Chengdu 610081;
3. Institute of Forest Ecology, Environment and Protection, CAF Beijing 100091;
4. Yanbian Forestry Bureau of Panzhihua City, Sichuan Province Yanbian 617002
林窗形成导致资源的再分配和微生境的异质性(Chazdon et al., 1991),影响种子进入、土壤种子库萌发、幼树生长、种内和种间竞争以及生态位分化等过程(宋新章等,2006)。Kneeshaw(1998)在研究中发现,在林窗大小一致但高度相差甚远的2个林窗中,由于太阳辐射不同,其他环境因子(如水分、温度、土壤营养物质等)也会有差异,这必然会使其中的植物个体在生理生化特性,甚至整个林窗更新上表现出差异。因此,可以用林窗长轴长度与边缘木高度的比值来表示林窗大小(谭辉等,2000),这个比值称为林窗开阔度,比值越大说明林窗的开阔程度越高。目前,林窗开阔度对林窗更新幼苗生长影响的研究已有报道(Zhu et al., 2003),王周平等(2000)和王开运(2004)研究了林窗开阔度的分布特征,但不同开阔度林窗的更新状况研究仍未涉及。
云南松(Pinus yunnanensis)系我国云贵高原特有树种,分布海拔为1 400~3 200 m, 属于喜光耐旱性树种,根系发达,适应性强,是川西南山地荒山造林的主要先锋树种和用材树种之一(杨玉坡等,1992)。在干热河谷上缘地带多与偏干性常绿阔叶树种形成以云南松为上层优势的异龄复层针阔混交林,目前,川西南山地云南松天然更新方面的研究已有报道(张旭东等,2006),但云南松林窗更新方面的研究不多(彭建松等,2005a;张旭东等,2006)。本研究分析林窗特征对川西南山地云南松更新的影响,探讨云南松林窗更新特征,为科学认识云南松林更新规律和林窗更新机制提供理论依据。
1 研究地概况研究地点位于川西南山地攀枝花市仁和区大黑山森林公园(101°41′ E,26°38′ N),海拔2 400~2 675 m,南坡,坡度为0~40°。母岩以花岗岩、片麻岩为主,土壤以山地黄棕壤为主。属南亚热带山地气候,光热充足,全年无霜期长,干湿季分明,年均气温15 ℃,≥10 ℃年积温4 200 ℃,年均降雨量1 200 mm,6—10月为雨季,占全年降雨量的90%左右。植被以川西南山地偏干性森林为主,主要建群种为云南松,混生有川滇高山栎(Quercus aquifolioides)、滇锥栎(Q. franchetii)及黄背栎(Q. pannosa)等,在海拔较高的地方存在少量的华山松(Pinus armandii)、滇油杉(Keteleeria evelyniana)。灌木层种类贫乏,以大白杜鹃(Rhododendron decorum)、南烛(Vaccinium bracteatum)为主。
2 研究方法 2.1 林窗年龄调查根据在当地林区工作多年的护林员的经验及林窗形成木(gap maker)的腐烂情况,估测林窗中最先倒下的一株形成木的死亡时间,推测出林窗年龄(臧润国,1999a)。
2.2 林窗干扰频率调查采用样线调查法,在云南松林中以某一随机点开始调查,由西向东方穿行,用皮尺记录走过的距离,遇见林窗时,调查林窗的形成年龄,并记录样线穿越林窗的长度。
2.3 林窗面积和开阔度调查采用样线调查法,以乌拉山庄为中心,向不同方向设立路线,仔细寻找沿途的每一个林窗,当发现林窗时,测量林冠林窗的长轴(L)和与长轴中心垂直的短轴(W);测量林窗边缘木的平均高度,作为林窗高度(H);调查林窗的形成年龄。根据所调查林窗接近椭圆形的特点,林窗面积(A)采用椭圆面积公式A=πLW/4计算(彭建松等,2005a)。林窗开阔度(O)用公式O=(L+W)/2/H计算(Zhu et al., 2003)。
2.4 林窗和林冠下更新调查由于调查区域在海拔2 000 m以上,土壤瘠薄,树种组成较为简单,云南松主要竞争对象为栎属(Quercus)树种。因此,将调查对象划分为3类:云南松、栎类和其他,其他类主要是群落演替过程中的杂木和灌木,调查其在林窗内的数量、地径和高度。共调查林窗65个,对其中的38个林窗及其相邻的云南松林中随机设置10 m×20 m的成对样方,调查样方内3类树种的数量、高度和地径。为了便于分析,文中按高度和地径将云南松划分为幼苗(H≤0.5 m)、幼树(H>0.5 m,地径≤15 cm)和成树(地径>15 cm)。
3 结果与分析 3.1 云南松林窗干扰频率运用样线法在云南松天然林中穿行了2 560 m,出现了109个林窗,林窗的线状密度为42.58个·km-1。在调查的2 560 m的线段上,处于林窗中的线段总长为676 m,所占比例平均为26.41%。在所调查的109个林窗中,年龄最大的林窗(50年)与年龄最小的林窗(< 1年)相差50年,则林窗的形成速率为0.85个·km-1a-1(42.58/50=0.85),每年约0.53%(26.41%/50=0.53%)的林分转化为林窗。因此,林窗干扰的间隔期为189年(1/0.53%=189)。
3.2 林窗和林冠下云南松更新比较云南松在林窗和林冠下表现出不同的更新特征(图 1)。林窗下幼苗的密度要显著高于林冠下(P < 0.05),但高度差异不大(P>0.05)。尽管云南松为阳性树种,对光照要求较高,但在云南松过熟林林下仍能萌发,这可能与过熟林林冠较稀疏有关。林窗中幼树的密度和平均高度明显大于林冠下的幼树(P < 0.05),表明林窗的发生有利于云南松幼树的生长和发育。林窗下成树密度明显高于林冠下(P < 0.05),但2者平均年龄差异显著(P < 0.05)。

|
图 1 林窗和林冠下不同生长阶段云南松种群特征 Figure 1 Characteristics of population of P. yunnanensis in different growth stages in gaps and canopys |
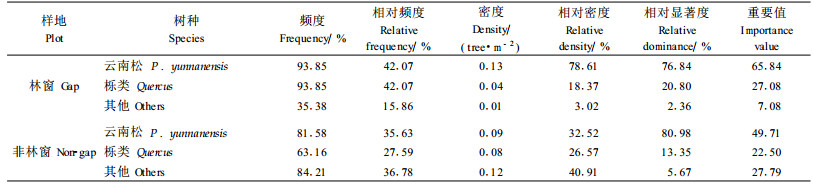
由表 1可知,无论在林窗内还是在非林窗中,云南松在重要值排序中均列第1位。林窗内云南松的频度、相对频度、密度和相对密度几项指标均大于非林窗,仅在相对显著度上低于非林窗的云南松。该结果表明,云南松并未因林窗干扰而影响其对资源的占据能力,过熟林林窗和非林窗中的云南松在更新群落中重要值相似。但在林窗中,栎类的重要值由非林窗下的第3位升至第2位,表明在林窗环境下,栎类生态位得到巩固和提高。
|
|
对年龄级为30~40年的27个林窗内的云南松密度进行分析,结果表明(图 2):云南松幼苗密度随林窗大小的增大迅速减小;林窗大小不同,幼树和成树密度不同,在面积为600~800 m2的林窗中密度较大;方差分析表明,不同大小林窗内云南松成树的平均地径差异不显著(P>0.05),平均高度差异显著(P < 0.05),幼树、幼苗的平均地径和高度的差异均显著(P < 0.05);大林窗下幼树和幼苗的个体较大,而成树在600 m2以上的林窗下平均高度最大。

|
图 2 不同面积林窗中云南松种群特征 Figure 2 Characteristics of regeneration population in P. yunnanensis in different gap areas |
对面积为200~400 m2的28个林窗内的云南松种群数量特征进行分析,结果表明(图 3):林窗中云南松种群的幼苗和幼树密度随林窗年龄增加而减小,而成树密度随林窗年龄增加呈上升趋势;方差分析表明,不同年龄林窗中云南松成树的地径和高度差异不显著(P>0.05),而幼苗和幼树差异显著(P < 0.05)。幼苗在年龄小于10年的林窗中个体最大,而幼树在年龄为20~30年的林窗中个体最大。

|
图 3 不同年龄林窗中云南松种群特征 Figure 3 Characteristics of P.yunnanensis regeneration population in gaps of different ages |
在调查中发现,在开阔度低于0.6的4个林窗中,只发现1株地径为16 cm的云南松,表明云南松在开阔度低于0.6的林窗中更新困难。对65个林窗内云南松种群数量的分析结果见图 4,在开阔度大于0.6的林窗下,云南松幼苗密度随开阔度增加而下降,而幼树和成树密度随开阔度的增加逐渐上升。方差分析表明,不同开阔度林窗内云南松成树的平均地径差异不显著(P>0.05),平均高度差异明显(P < 0.05),幼树、幼苗的平均地径和高度差异显著(P < 0.05)。从图 4可知,林窗开阔度越大,林窗内云南松个体越大。

|
图 4 不同开阔度林窗中云南松种群特征 Figure 4 Characteristics of P. yunnanensis regeneration population in gaps of different canopy openness |
为进一步探讨林窗下云南松的更新特征,将云南松按高度和地径分为6级(Ⅰ:H≤0.5 m; Ⅱ:H>0.5 m,地径(DGL)≤5 cm; Ⅲ: 5 cm < DGL≤10 cm; Ⅳ: 10 cm < DGL≤15 cm; Ⅴ:15 cm < DGL≤20 cm; Ⅵ:DGL>20 cm),分析其数量特征(图 5)。在小林窗(< 200 m2)、幼年林窗(< 10年)和开阔度为0.6~1.0的林窗中,Ⅰ级云南松(即云南松幼苗)数量最高。随着林窗内大径级云南松数量的增多,林窗内竞争加剧,导致云南松幼苗数量逐渐降低。在年龄 < 10年的林窗中不存在DGL>20 cm的云南松。在开阔度 < 0.6的林窗中云南松几乎不存在更新。随着开阔度的增大,Ⅰ,Ⅱ级云南松数量逐渐降低,Ⅲ,Ⅳ,Ⅴ及Ⅵ级云南松数量逐渐增大,表现出较为规律的变化。从总体上看,不同面积、年龄和开阔度的林窗下Ⅲ级云南松与Ⅳ级云南松数量存在明显差异,Ⅳ级云南松的数量平均仅为Ⅲ级云南松的2/5。该结果表明,Ⅲ级云南松向Ⅵ级云南松转化的过程中出现了较为明显的自疏,是云南松更新的一个关键阶段。

|
图 5 林窗内不同大小级云南松数量特征 Figure 5 Number of different sizes P. yunnanensis in gaps |
研究结果表明,对阳性树种云南松而言,林窗干扰对林窗更新的幼苗和幼树生长有促进作用。同时,林窗干扰并未影响云南松对资源的占据能力。因此,林窗干扰是云南松维持天然更新的有效途径。Zhu等(2005)对阳性树种樟子松的天然更新研究中发现, 其更新苗具有一定的耐荫性, 但如果没有较大林窗或较大的其他干扰, 更新苗木将很难进行入主林冠层。云南松的天然更新同样表现出类似的特点。
在年龄级或大小级相同的林窗中讨论云南松林窗更新,更能体现云南松林窗更新的规律。本次研究发现,相同年龄级的林窗中,不同面积林窗下云南松密度随林窗增大而降低;相同大小级的林窗中,随着林窗形成时间延长云南松密度逐渐降低。该结果和许多研究结果一致(臧润国等,1999b;刘庆等,2002;罗大庆等,2002;陶建平等,2004)。
开阔度作为林窗特征之一,能很好地反映林窗对云南松更新的影响。本次研究表明,云南松在较小的林窗(开阔度小于0.6)中不存在更新,而在大林窗(开阔度大于1.5)中更新最好,同时随着开阔度的增大,不同大小的云南松更新表现出较为规律的变化。可见,开阔度综合考虑了林窗高度和面积大小,是体现林窗大小的一个综合指标。
林窗大小、形成时间和开阔度不同,更新种群的数量动态不同。本研究发现,云南松幼苗(高度 < 0.5 m)在小林窗、幼年林窗以及开阔度低的林窗中密度最高,呈现“只见幼苗,不见大树”的景象,这表明幼苗在该类林窗的林窗中萌发率高,但死亡率也较高。在面积较大和开阔度高的林窗中,大个体云南松的密度较高。随着林窗面积和开阔度增加,光的有效性增加(Zhu et al., 2003),来自大树对水分和营养的竞争减少,有利于林窗更新(Madsen et al.,1997)。
林窗中地径为5~10与10~15 cm的云南松数量差距最为明显,表明在这一时期云南松表现出激烈的种内竞争,自疏现象明显。一方面,在竞争中占据优势的云南松出现较快的生长,另一方面,在竞争中处于劣势的云南松生长停滞甚至死亡。因此,地径在10~15 cm的云南松是林窗更新的关键,对云南松林窗更新产生重大影响。这一阶段云南松出现的分化,可能造成云南松年龄结构断代,最终导致云南松林的衰退,在以后的森林经营中应适当的加以人工抚育。
云南松是偏干性森林的阳性树种,在适宜的地形、时间和干扰影响条件下,云南松会成为偏途顶级群落(杨玉坡等,1992)。在实施天然林保护工程以前,当地居民常割取松脂用于引火和照明,加上大黑山土壤瘠薄,海拔较高,坡度大,造成云南松过早死亡形成林窗。过去的研究表明,从温带→亚热带→热带,林窗干扰返回间隔期依次缩短,森林循环速率呈递增趋势(臧润国等,1999c;刘金福等,2003;杨修,2002;王开运,2004;谭辉等,2007)。本次研究发现,大黑山云南松林林窗干扰间隔期为189年,与云南白马河林场的云南松林的400年相差较大(彭建松等,2005b),更接近于海南热带山地雨林的160年(臧润国等,1999a),表现出较快的循环速率。这种快速的循环,可能是云南松成为该区域偏途顶级群落的重要原因之一。
刘金福, 洪伟, 李俊清, 等. 2003. 格氏栲林林窗自然干扰规律. 生态学报, 23(10): 1991-1999. DOI:10.3321/j.issn:1000-0933.2003.10.006 |
刘庆, 吴彦. 2002. 滇西北亚高山针叶林林窗大小与更新的初步分析. 应用与环境生物学报, 8(5): 453-459. DOI:10.3321/j.issn:1006-687X.2002.05.002 |
罗大庆, 郭泉水, 薛会英. 2002. 西藏色季拉山冷杉原始林林隙更新研究. 林业科学研究, 15(5): 564-569. DOI:10.3321/j.issn:1001-1498.2002.05.010 |
彭建松, 柴勇, 孟广涛, 等. 2005a. 云南金沙江流域云南松天然林林隙特征. 浙江林学院学报, 22(1): 50-55. |
彭建松, 柴勇, 孟广涛, 等. 2005b. 云南金沙江流域云南松天然林林隙更新研究. 西北林学院学报, 20(2): 114-117. |
宋新章, 肖文发. 2006. 林隙微生境及更新研究进展. 林业科学, 42(5): 114-119. |
谭辉, 朱教君, 康宏樟, 等. 2007. 林窗干扰研究. 生态学杂志, 26(4): 587-594. DOI:10.3321/j.issn:1000-4890.2007.04.025 |
陶建平, 臧润国. 2004. 海南霸王岭热带山地雨林林隙幼苗库动态规律研究. 林业科学, 40(3): 33-38. DOI:10.3321/j.issn:1001-7488.2004.03.005 |
王开运. 2004. 川西亚高山森林群落生态系统过程. 四川: 四川科技出版社.
|
王周平, 李旭光, 石胜友, 等. 2000. 缙云山森林林隙形成特征的研究. 西南师范大学学报:自然科学版, 25(3): 305-309. |
杨修. 2002. 长白山暗针叶林林隙一般特征及干扰状况. 生态学报, 22(11): 1825-1831. DOI:10.3321/j.issn:1000-0933.2002.11.005 |
杨玉坡, 李承彪. 1992. 四川森林. 北京: 中国林业出版社.
|
臧润国, 刘静艳, 董大方. 1999a. 林隙动态与森林生物多样性. 北京: 中国林业出版社.
|
臧润国, 徐化成, 高文韬. 1999b. 红松阔叶树林主要树种对林隙大小及其发育阶段更新反应规律的研究. 林业科学, 35(3): 2-7. |
臧润国, 余世孝, 刘静艳, 等. 1999c. 海南霸王岭热带山地雨林林隙更新规律的研究. 生态学报, 19(12): 152-158. |
张旭东, 费世民, 周金星. 2006. 中国西部山地生态脆弱区植被恢复研究. 北京: 科学出版社.
|
Chazdon R L, Pearcy R W. 1991. The importance of sunflecks for forest understory plants. Biology Science, 41(11): 760-766. |
Kneeshaw D D. 1998. Canopy gap characteristics and tree replacement in the southeastern boreal forest. Ecology, 79(3): 789-794. |
Madsen P, Larsen J B. 1997. Natural regeneration of beech (Fagus sylvatica) with respect to canopy density, soil moisture and soil carbon content. F orest Ecology and Management, 97: 103-113. |
Zhu Jiaojun, Kang Hongzhang, Tan Hui, et al. 2005. Regeneration characteristics of natural Mongolia Pine(Pinus slvestris var. mongolica) forests on sandy land in Honghuaerji. Journal of Forestry Research, 16(4): 253-259. DOI:10.1007/BF02858184 |
Zhu Jiaojun, Matsuzaki T, Lee Fengqin, et al. 2003. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. Forest Ecology and Management, 182: 339-354. DOI:10.1016/S0378-1127(03)00094-X |