 2008, Vol. 44
2008, Vol. 44文章信息
- 梁军, 王媛, 贾秀贞, 张星耀.
- Liang Jun, Wang Yuan, Jia Xiuzhen, Zhang Xingyao
- 溃疡病菌对杨树愈伤组织细胞膜透性、可溶性糖及MDA含量的影响
- Effects of Infection with Botryosphaeria dothidea on Cell Membrane Permeability, Soluble Sugar and MDA Content in Poplar Calli
- 林业科学, 2008, 44(8): 72-77.
- Scientia Silvae Sinicae, 2008, 44(8): 72-77.
-
文章历史
- 收稿日期:2007-04-20
-
作者相关文章



杨树溃疡病是我国杨树人工林重大生物灾害之一,严重制约了杨树人工林的发展(张星耀, 2003)。该病在我国东北、华北、华东和西北等地均普遍发生,造成育苗和造林的失败,经济和生态损失惊人,严重地区林木的发病率高达96.1%~100%,感病指数达68.7%~95.8%(苏晓华等, 2003; 黄烈健等, 2003)。自杨树溃疡病1955年在北京首次被发现并报道以来,许多学者就其病原学、病理学、流行学和防治学等进行了研究,取得了一系列研究成果,但在杨树与溃疡病菌互作基础方面的研究仍较少。本研究小组致力于溃疡病病原真菌类群的分子遗传多样性及溃疡病害的生态控制技术研究(张星耀等, 1999; 2000;梁军等, 2005),而寄主与溃疡病菌互作是揭示病害发生和寄主抗性机制的重要内容(刘会香等, 2005)。
植物对病原微生物的防卫反应主要包括植物细胞对病原菌的识别、胞内信号的转换与传导、防卫反应的开启与系统获得抗性(system acquired resistance, SAR)的形成等(周建明等, 1999)。其中早期互作中细胞膜透性、可溶性糖及丙二醛(MDA)含量的变化互相有联系,并与树种抗病性有一定的相关性,可作为筛选抗病种及进行药剂调控与保护的研究靶标(郑翠明等, 1998; 郑殿峰等, 2004),是相关研究中的重点,同时可从代谢变化的角度揭示寄主与病原菌早期识别、侵染及表现症状的互作机制。在农作物上关于寄主与病原菌互作中膜脂过氧化及保护酶系统的变化报道很多,但在杨树与溃疡病菌互作中相关研究则较少。本文选择了2种抗性水平不同的杨树树种,研究其愈伤组织在接种溃疡病菌后细胞膜透性、可溶性糖及MDA含量的变化及与树种抗性的关系,阐述寄主与病原菌之间的互作,为进一步明确和揭示溃疡病菌对杨树的作用机理提供依据。
1 材料与方法 1.1 供试材料 1.1.1 供试植物材料毛白杨(Populus tomentosa),高抗树种,自然状态下感病指数0~6.0;北京杨(P. beijingensis),高感树种,自然状态下感病指数45.1以上(王颖等, 1996)。2006年2月,选取毛白杨、北京杨1年生健康枝条,取树皮组织(带形成层)为外植体,经自来水冲洗拭干后,无菌条件下对材料进行表面消毒,然后剪成0.5 cm×0.5 cm的小块,接种于MS+2, 4-D 0.5 mg·L-1+6-BA 2 mg·L-1诱导培养基上,诱导产生愈伤组织后继代培养,建立愈伤组织无性系(赵华燕等, 2001)。继代培养2~3次后,选择质地紧密、大小均一、1.2 mm×1.2 mm左右的愈伤组织进行溃疡病菌的接种试验。
1.1.2 供试病原菌选择本研究室分离并保存的杨树溃疡病病原(Botryosphaeria dothidea)菌株,编号为CXY160(致病力测定为中等)。将菌株在27℃条件下于PDA平板上暗培养7 d,无菌条件下用灭菌打孔器在菌落边缘打取直径为5 mm的菌饼作为接种体备用。
1.2 接种及取样方法 1.2.1 接种方法无菌条件下选择大小均一的愈伤组织块置于盛有2%水琼脂的培养皿中,每皿放置5块,将菌饼接种于愈伤组织块上,对照组接种同样大小的灭菌PDA,放于培养室中25℃黑暗培养。每树种设接种和对照2个组,每组50块愈伤组织。
1.2.2 取样方法取样时用打孔器从中央打取直径8 mm的圆柱体,在上端切取圆片,称取0.5 g作为样品,迅速投入液氮中,并置于-80℃保存,待样品全部获取完毕后同时测定各项指标。取样时采用完全随机方法,每次取5个组织块的混合样,从混合样中随机称取3个样品作为重复。
1.3 生理生化指标测定 1.3.1 愈伤组织受害程度接菌后,分别于0、12、24、48、72、96、120、144 h观察记录菌丝生长情况及愈伤组织变化情况,参照张立钦等(1989)的分级标准并根据本试验情况,将愈伤组织接种病原菌后的感病程度分为5级(表 1),用感病指数来表示愈伤组织受害程度。

|
|
|

用相对电导率表示膜透性。分别于0、24、48、72、96、120、144 h取样,用DDS-307型电导仪测定相对电导率,将样品置于20 mL重蒸水中,真空(气压1×10-3Pa)抽气15 min后测定电导率为初电导率,然后将各材料煮沸10 min,冷却至室温后再测电导率为终电导率,初电导率与终电导率之比为相对电导率。
1.3.3 可溶性糖含量测定取样时间同1.3.2。采用蒽酮比色法测定,分析纯蔗糖作标准曲线(黄顶等, 2006),单位为μg·g-1FW。
1.3.4 MDA含量测定取样时间同1.3.1。硫代巴比妥酸比色法测定(周琦, 1993),以μmol·g-1 FW表示。
2 结果与分析 2.1 溃疡病菌对愈伤组织的损伤无论是抗病种毛白杨还是感病种北京杨,接菌后12 h内愈伤组织上均没有肉眼可观察到的菌丝生长,只在接种的菌丝块外缘出现菌丝膨大和少量生长(200倍显微镜观察,未显示图像);12~24 h,首先在感病种愈伤组织接种点周围发现菌丝生长,抗病种上的菌丝生长则平均滞后5 h以上,此时菌丝的生长局限于接菌点周围,扩展不明显;24 h后菌丝扩展加速,至72 h,2树种组织块已大部分或全部被菌丝所覆盖,接菌点周围开始出现不同程度的褐变;144 h时,大部分组织块均已严重褐变,并呈现水渍状、腐烂和皱缩(图 1)。感病指数的变化与愈伤组织受害情况吻合,表现为接菌后呈现上升趋势,感病种上升幅度始终高于抗病种(表 2)。

|
图 1 不同时间溃疡病菌侵染对杨树愈伤组织的影响 Figure 1 Effect of B. dothidea on poplar callus at different time after inoculation a-d:毛白杨愈伤组织. a:未接菌; b:接菌48 h, 接菌点开始有少量菌丝生长; c:接菌72 h, 接菌点周围轻微褐变; d:接菌144 h, 完全变褐, 并呈现腐烂状; e-h:北京杨愈伤组织. e:未接菌; f:接菌48 h, 菌丝快速扩展, 菌丝下组织有轻微褐变; g:接菌72 h, 严重褐变, 接菌点处开始呈现腐烂状; h:接菌144 h, 腐烂严重, 组织块皱缩变形。 a-d: P. tomentosa callus. a: untreated; b: 48 h after inoculation (hai), little hyphae around inoculation site; c: 72 hai, slight brownness; d: 144 hai; e-h: P. beijingensis callus. e: untreated; f: 48 hai, hyphae develop rapidly, but no clear brownness in inoculation site; g: 72 hai, serious brownness and rust; h: 144 hai, heavy rust and shrinking. Bar=3 mm. |
|
|
从愈伤组织的褐变现象看,2树种褐变的出现时间、程度及发展速度有较明显的差别。感病种出现褐变后,扩展速度和严重程度高于抗病种。接菌48 h时,抗病种接菌点处有轻微褐变,并局限于小范围内,120 h时褐变明显,组织逐渐变为褐色,而对照愈伤组织仍为淡黄色;感病种接菌48 h后也开始出现褐变,褐变速度比抗病种要快,120 h时愈伤组织块上已有大面积严重褐变, 而对照愈伤组织未发现菌丝生长,无褐变现象;接菌144 h时抗病种和感病种愈伤组织均出现较为严重的腐烂,感病种愈伤组织皱缩变形。
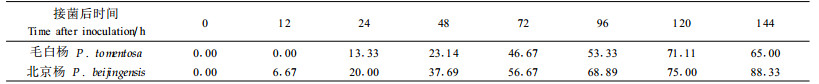
2.2 溃疡病菌对愈伤组织细胞膜透性的影响细胞膜透性(以相对电导率表示)变化见表 3。结果显示,24 h内2树种相对电导率在处理和对照中变化均不明显,随接菌时间的延长,病原菌对愈伤组织细胞膜透性的破坏作用逐渐加大,表现为相对电导率持续增加,并且2树种有相似的变化趋势,只是增加幅度不同。接菌后48~144 h,抗病种相对电导率由38.33%增至112.67%,增长幅度为-5.89%~57.06%,而感病种相对电导率由74.17%增至167.67%,增长幅度为70.62%~96.91%,明显高于抗病种。北京杨48 h后相对电导率开始大幅上升,至后期(120~144 h)升幅更大,并于144 h达到细胞膜透性增加的高峰,此时相对电导率为167.67%,比相应对照增加94.45%;而毛白杨相对电导率从72 h才开始大幅上升,比北京杨滞后24 h,上升幅度也远小于感病种。方差分析表明:毛白杨从接菌0~72 h,北京杨从0~48 h,处理与对照的差异不显著(P>0.05),而随后毛白杨和北京杨的处理与对照相比均在0.05水平上差异显著(t=2.292,P=0.031<0.05)。结合图 1所示,北京杨在接菌72 h时菌丝已长满愈伤组织,褐变明显,至144 h时整个愈伤组织块已呈现严重的腐烂和皱缩,而抗病种整个变化过程滞后于感病种,说明膜透性的变化与愈伤组织受害程度呈正相关,与抗病性呈负相关。
|
|
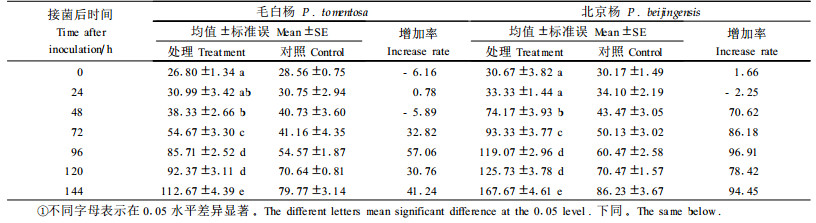
杨树愈伤组织接菌后可溶性糖含量变化如表 4所示。从表 4中看出,接菌后,毛白杨对照可溶性糖含量在0~72 h有小幅下降,随后变化不大,处理在48 h时略高于对照,为24.96 μg·g-1FW,增加幅度只有11.43%,其余时间则低于对照,方差分析结果显示t=-1.062,P=0.295>0.05,处理与对照相比差异不显著;北京杨愈伤组织24 h时可溶性糖含量达到高峰,为49.42 μg·g-1FW,比相应对照增加了33.50%,随后下降至接近对照,但比对照略高,后期(120 h)又有一个小高峰,比相应对照增加28.55%,方差分析结果显示t=2.71,P=0.010<0.05,处理与对照相比差异显著。
|
|

在黑杨上,抗褐斑病无性系叶部还原糖和可溶性糖含量较感病无性系低且呈下降趋势,感病无性系则呈上升趋势(万贤崇等, 1995)。病原侵染寄主时大多先分泌各种细胞壁水解酶导致可溶性糖积累,植物体内高的含糖量为病原的繁殖及病原分解植物体内木质素、纤维素及酚类化合物提供能源,促进病原对这些物质的大量分解(Entry et al., 1991; 1993),导致病害症状的出现。本研究中杨树愈伤组织接菌后,感病种可溶性糖含量上升趋势大于抗病种。
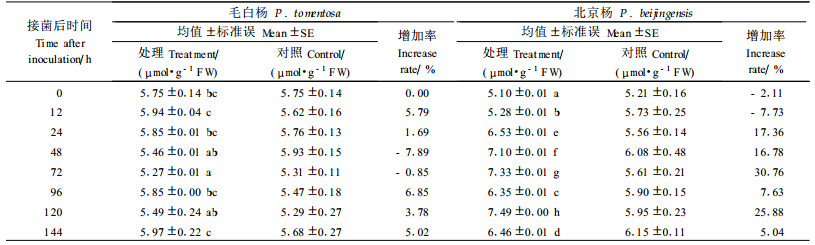
2.4 溃疡病菌对愈伤组织MDA含量的影响表 5表明:2树种对照的MDA含量在不同时间上的差异不显著,接菌后不同时间在抗病和感病种上则变化差异较大。毛白杨MDA含量24 h内与对照相差不大,24 h后趋于下降,96 h后开始上升,96~144 h间始终小幅高于对照,说明前期细胞的膜脂过氧化程度不明显,后期则略有加重,但没有发生严重的膜脂过氧化;北京杨则从24 h开始上升,并在72 h和120 h达到高峰,MDA含量分别为7.33 μmol·g-1FW和7.49 μmol·g-1FW,比相应对照增加30.66%和25.88%,远大于毛白杨的增加幅度。从结果来看,毛白杨处理比对照的增减比率的变化幅度为-7.89%~6.85%,明显低于北京杨的变化幅度-7.73%~30.76%,说明抗病种受病菌侵染后质膜电解质外渗率变幅较小,抗病种能有效地减少这种变幅,使细胞变化趋于稳定。毛白杨膜脂过氧化程度小于北京杨,从细胞膜角度揭示了毛白杨抗病性强于北京杨。
|
|
溃疡病菌侵染后,2树种愈伤组织细胞膜透性上升,相对电导率显著增加,且感病种上升幅度大于抗病种。因此,细胞膜透性的变化情况反映了植物抗病性的强弱。
溃疡病菌接种愈伤组织24 h后菌丝开始不同程度的生长,72 h后逐渐覆盖整个愈伤组织,愈伤组织颜色加深,最后变为褐色,同时出现水渍化,变化的不同时期细胞相对电导率的结果说明细胞膜受到严重伤害。感病种北京杨愈伤组织褐变的速度快,48 h时已有部分愈伤组织接种点周围出现轻微褐变,而抗病种大多在72 h后才开始褐变,显示出了抗感树种在溃疡病菌作用下不同的保护性反应,也说明抗病种愈伤组织细胞在病菌胁迫下所受的损伤小于感病种,细胞膜透性与寄主抗病性呈负相关。
3.2 病菌侵染与可溶性糖含量的变化可溶性糖含量的变化在抗/感病种间变化差异较大,感病种可溶性糖含量呈上升趋势,在24 h和120 h达到高峰,抗病种可溶性糖含量48 h时高于对照,但比相应对照增加幅度小,其余测定时间低于对照,方差分析表明抗病种处理与对照相比可溶性糖含量差异不显著。
本研究中感病种正常水平的可溶性糖含量高于抗病种。分析溃疡病菌在感病种愈伤组织上的生长状况,发现24 h时可溶性糖含量达到最高,正好是菌丝开始生长之时,由于菌丝生长导致组织水解加强,光合产物输出减慢,造成糖类堆积。接种后期则随着病菌的进一步扩展,细胞遭到破坏,内容物加速渗漏,导致可溶性糖含量的第2次升高,可溶性糖含量变化与菌丝生长有一定的相关性。大量的研究表明:生理、生化指标的大小和活性高低在抗病和感病种上的表现并不呈现一致规律,有些病害系统中所得的结果甚至矛盾(Gibbs et al., 1972; Vidhyasekaran et al., 1974)。而植物在受到病原菌侵染后,迅速启动生理、生化指标的变化并在短时间内调节这些指标恢复至正常状态的能力或许才是判断寄主植物是否抗病的更科学依据。
3.3 细胞膜脂过氧化与抗病性的关系接菌后不同时间的感病种和抗病种的MDA含量变化差异较大。感病种北京杨MDA含量的变化幅度为-7.73%~30.76%,明显高于抗病种毛白杨的-7.89%~6.85%变化幅度。
MDA的大量积累能交联脂类、核酸、糖类和蛋白质,从而进一步对质膜的结构和功能造成不良影响,导致膜结构改变,影响膜的流动性及酶的结合力,最终由于代谢紊乱而抑制植物的生长(潘华珍等, 1984)。研究表明:遭到病原菌侵染后MDA在短期内含量急剧上升,说明机体自身代谢功能特别是保护酶系统的调节失去平衡,导致活性氧过剩,膜脂过氧化作用加强,但随后保护酶的活性又趋于稳定或不再急剧下降,MDA含量不再持续增加,可能植物体内一些应激蛋白开始起调节作用(Lindquist et al., 1988)。本研究发现,接种病菌后,感病种MDA含量持续上升,尤其在24~72 h间与相应对照差别较大,这一时期也是菌丝大量生长并对组织造成严重影响的时期,细胞相对电导率开始大幅上升,说明发生了较为严重的膜脂过氧化,随后相对电导率增加更快,细胞丧失了自我恢复能力,最终导致死亡。抗病种MDA含量变化则趋于平稳,没有发生明显的膜脂过氧化。
黄顶, 王堃. 2006. 典型草原常见牧草春季萌动期可溶性糖及内源激素动态研究. 应用生态学报, 17(2): 210-214. DOI:10.3321/j.issn:1001-9332.2006.02.010 |
黄烈健, 苏晓华. 2003. 我国杨树溃疡病研究进展. 世界林业研究, 16(4): 49-53. DOI:10.3969/j.issn.1001-4241.2003.04.010 |
李海英, 刘亚光, 杨庆凯. 2002. 大豆树种感染灰斑病前后可溶性糖含量的比较. 中国油料作物学报, 24(3): 50-51. DOI:10.3321/j.issn:1007-9084.2002.03.013 |
梁军, 姜俊清, 刘会香, 等. 2005. 我国杨树与溃疡病菌互作的病理学研究. 林业科学研究, 18(2): 214-221. DOI:10.3321/j.issn:1001-1498.2005.02.022 |
刘会香, 贾秀贞, 吕全, 等. 2005. 中国杨树溃疡病的发生与防治. 世界林业研究, 18(4): 60-63. DOI:10.3969/j.issn.1001-4241.2005.04.012 |
潘华珍, 冯立明, 许彩民, 等. 1984. 丙二醛对红细胞的作用. 生物化学与生物物理进展, (2): 34-37. |
苏晓华, 张冰玉, 黄秦军, 等. 2003. 我国林木基因工程研究进展和关键领域. 林业科学, 39(5): 111-118. DOI:10.3321/j.issn:1001-7488.2003.05.017 |
万贤崇, 汪安琳. 1995. 杨树糖代谢与褐斑病发生的关系. 南京林业大学学报, 19(1): 37-40. |
王颖, 胡景江, 朱玮, 等. 1996. 杨树溃疡病寄主诱导抗病性的研究. 西北林学院学报, 11(1): 1-4. |
张立钦, 李传道, 黄敏仁. 1989. 杨树组织培养愈伤组织对水疱型溃疡病的抗性. 南京林业大学学报, 13(4): 9-15. |
张星耀, 骆有庆. 2003. 中国森林重大生物灾害. 北京: 中国林业出版社.
|
张星耀, 赵仕光, 吕全, 等. 2000. 树木溃疡病病原真菌类群分子遗传多样性研究Ⅱ:Botryosphaeria属28S rDNA-PCR-RFLP和RAPD解析. 林业科学, 36(2): 75-81. DOI:10.3321/j.issn:1001-7488.2000.02.012 |
张星耀, 赵仕光, 朴春根, 等. 1999. 树木溃疡病病原真菌类群分子遗传多样性研究Ⅰ:小穴壳菌属、疡壳孢属、壳囊孢属、盾壳霉属分类地位的分子证明. 林业科学, 35(3): 34-40. DOI:10.3321/j.issn:1001-7488.1999.03.007 |
赵华燕, 卢善发, 晁瑞堂. 2001. 杨树的组织培养及其基因工程研究. 植物学通报, 18(2): 169-176. DOI:10.3969/j.issn.1674-3466.2001.02.007 |
郑翠明, 滕冰, 高风兰, 等. 1998. 不同种粒抗性大豆树种感染SMV后可溶性蛋白和游离氨基酸的研究. 植物病理学报, 28(3): 227-231. DOI:10.3321/j.issn:0412-0914.1998.03.008 |
郑殿峰, 梁喜龙, 左豫虎, 等. 2004. 大豆根腐病菌对大豆幼苗生理生化指标的影响. 中国油料作物学报, 26(3): 57-61. DOI:10.3321/j.issn:1007-9084.2004.03.013 |
周建明, 朱群, 白永延. 1999. 高等植物防卫反应的信号传导. 植物学通报, 16(3): 228-237. DOI:10.3969/j.issn.1674-3466.1999.03.005 |
周琦. 1993. 植物生理生化试验指导. 北京: 中国农业出版社.
|
Entry J A, Cromack K Jr, Kelsey R G, et al. 1991. Response of Douglas-fir to infection by Armillaria ostoyae after thinning or thinning plus fertilization. Phytopathology, 81(6): 682-689. DOI:10.1094/Phyto-81-682 |
Entry J A, Donnelly P K, Cromack K J R. 1993. Effect of nitrogen and carbon sources on lignin and cellulose degradation by Armillaria ostoyae. Forest Pathology, 23(3): 129-137. DOI:10.1111/efp.1993.23.issue-3 |
Gibbs A F, Varma A, Woods R D. 1972. Effect of sugar content of Poapratensis on Helminthosporium leaf spot. Physiological Plant Pathology, 2: 279-287. DOI:10.1016/0048-4059(72)90011-2 |
Lindquist S, Craig E A. 1988. The heat-shock proteins. Annu Rev Genet, 22: 631-677. DOI:10.1146/annurev.ge.22.120188.003215 |
Vidhyasekaran D. 1974. Possible role of sugars in restriction of lesion development in finger millet leaves infected with Helminthosporium tetrmera. Physiological Plant Pathology, 4: 457-467. DOI:10.1016/0048-4059(74)90031-9 |