 2008, Vol. 44
2008, Vol. 44文章信息
- 王彦平, 李静, 杨敏生, 梁海永.
- Wang Yanping, Li Jing, Yang Minsheng, Liang Haiyong
- 转不同抗虫基因741杨的抗虫选择性
- Insect-Resistance Selectivity of Transgenic Hybrid Poplar 741
- 林业科学, 2008, 44(8): 67-71.
- Scientia Silvae Sinicae, 2008, 44(8): 67-71.
-
文章历史
- 收稿日期:2007-03-07
-
作者相关文章



近年来,随着植物抗虫基因工程的研究及其快速发展,转基因植物的合理利用问题受到了广泛关注。目前,在杨树抗虫基因工程育种方面国内外已经做了大量工作,人们构建、改建了多种基因并将其成功转入杨树(苏晓华等,2003;王敏杰等,2002;王学聘等,1997;郭同斌等,2004;田颖川等,2000;Zhang et al., 2005;Van Frankenhuyzen et al., 2004;Balestrazzi et al., 2006)。其中苏云金芽孢杆菌(Bacillus thuringiensis, 简称Bt)抗虫基因的研究最多,发展也最快。根据其编码的杀虫蛋白的杀虫特性,可将其粗分为6大类(李海涛等,2004)。目前,在杨树抗虫基因工程中应用最多的是抗鳞翅目害虫的Btcry1和抗鞘翅目害虫的Btcry3类基因,转不同类型抗虫基因杨树对靶标害虫均表现出一定的抗虫性。为了深入了解转抗虫基因植物的抗虫选择性, 为转抗虫基因植物的合理利用提供依据,本文对转不同类型Bt抗虫基因741杨的抗虫性进行了研究。
1 材料与方法 1.1 材料参试的株系为转BtCry3A基因741杨的7个株系和转双抗虫基因(BtCrylAc+API) 741杨的2个株系PB29和PB17。以苗圃中生长的转基因741杨各株系和未转基因741杨1年生植株为试验材料。
测试昆虫为人工饲养的鞘翅目叶甲科的柳蓝叶甲(Plagiodera versicolora)和鳞翅目舟蛾科的杨扇舟蛾(Clostera anachoreta)。
1.2 方法 1.2.1 转基因741杨外源基因的分子生物学检测杨树DNA的制备采用CTAB法(王关林等,2002)。采用PCR方法检测外源基因。BtCrylAc基因引物序列:5′CTG ACG TAA GGA TGA CGC AC3′,5′ACT ATT GAT AGT CGC GGC ATC3′。PCR扩增程序:94℃ 50 s, 55℃ 60 s, 72℃ 100 s, 共30个循环。BtCry3A基因引物序列:5′ACC GTC TCT GGT AAG CTC GGT CTT3′,5′TGG CCA AGC GAG GAC CCC TGG AAG 3′。PCR扩增程序:94 ℃、50 s, 52℃、60 s, 72℃、60 s, 共30个循环。反应结束后扩增产物在1%琼脂糖凝胶上电泳检测。
1.2.2 转抗虫基因741杨Bt毒蛋白的检测采用ELISA法测定。操作步骤按美国Agdia公司生产的BtCrylAb/CrylAc和BtCry3A ELISA平板试剂盒说明进行。测定转BtCry3A基因741杨和转双抗虫基因(BtCrylAc+API) 741杨不同株系毒蛋白含量,以未转基因741杨为对照,计算相对含量。
1.2.3 转基因741杨抗虫性检测1) 柳蓝叶甲 由室外捕捉柳蓝叶甲成虫于室内用普通741杨叶片在瓶中饲养,待成虫产卵于叶片表面,将卵块用剪刀剪下,放到培养皿(培养皿内壁上贴有潮湿的滤纸)中孵化。大约3~4 d卵孵化为幼虫,将幼虫分装到放有转抗虫基因杨树叶片的培养皿中进行虫试。每个培养皿中饲养20~30头幼虫,每个株系设3个重复,每天记录死亡个数。计算每天的累计死亡率。2)杨扇舟蛾 待杨扇舟蛾卵块孵化后,随机地从中挑选幼虫,用毛笔轻移到清洁干净的高为12 cm、直径8.5 cm的罐头瓶中,每瓶10头左右,4次重复,放入新鲜的转抗虫基因(BtCry3A) 741杨当年生叶片,然后用带通气孔塑料薄膜扎紧瓶口。每2 d换一次新鲜叶片,记录幼虫的死亡情况。总死亡率=饲养末期的死亡总数/初期的饲养总数×100%。
2 结果与分析 2.1 不同外源Bt基因的PCR扩增将转抗虫基因(BtCry1Ac+API基因)741杨的2个株系和转抗虫基因(BtCry3A)741杨的6个株系和分别提取叶片基因组总DNA,分别以质粒pBtiA和pBCC3作为阳性对照,未转基因植株作为阳性对照,用BtCry1Ac和BtCry3A基因序列设计引物,进行PCR扩增。扩增结果如图 1、2所示。

|
图 1 BtCry1Ac基因检测结果 Figure 1 Test for BtCry1Ac gene 泳道3 Lane3:marker; +:阳性对照Positive control; -:阴性对照Negative control; 泳道1-2 Lane 1-2:转基因(BtCry1A+API)741杨株系Subclones of transgenic poplar clone 741 carrying BtCry1A +API genes PB29, PB17. |

|
图 2 BtCry3A基因检测基因 Figure 2 Test for BtCry3A gene 泳道7 Lane7:marker; +:阳性对照Positive control; -:阴性对照Negative control; 泳道1-6 Lane 1-6:转抗虫基因(BtCry3A)741杨株系Subclones of transgenic poplar clone 741 carrying BtCry3A gene CC84、CC31、CC71、CC70、CC53、CC11. |
由图 1、2可以看出,转BtCry1Ac+API基因741杨的2个株系经PCR扩增后,作为阳性对照的pBtiA扩增出1条750 bp的片段,转基因的各株系经PCR扩增后,均得到了1条与预期长度同样大小的750 bp的片段,而未转基因植株未出现PCR扩增特异条带。同时,转抗虫基因(BtCry3A)741杨各株系经PCR扩增后,作为阳性对照的pBCC3扩增出1条612 bp的片段,转基因的各株系经PCR扩增后,均得到了1条与预期长度同样大小的612 bp的片段,未转基因植株没有出现PCR扩增片段。电泳分析结果表明,2种Bt抗虫基因分别在转基因741杨不同株系中稳定存在。
2.2 不同外源Bt基因毒蛋白检测利用美国Agdia公司生产的BtCrylAb/CrylAc和BtCry3A ELISA平板试剂盒,分别测定转不同Bt基因741杨叶片中Bt毒蛋白含量,结果见图 3、4。

|
图 3 转抗虫基因(BtCry1Ac+API)741杨叶片毒蛋白的含量 Figure 3 The toxin protein concentration in leaf of transgenic hybrid poplar clone 741 carrying BtCry1Ac+API gene |

|
图 4 转抗虫基因(BtCry3A)741杨叶片毒蛋白的含量 Figure 4 The toxin protein concentration in leaf of transgenic hybrid poplar clone 741 carrying BtCry3A gene |
从图 3可见,2个参试的转(BtCry1Ac+API)基因株系中,BtCry1型的毒蛋白含量分别为0.012 7%和0.009 9%。高抗虫性株系pb29的毒蛋白含量高于中等抗虫性的株系pb17。图 4结果表明:5个转BtCry3A基因的株系中,BtCry3型毒蛋白含量为0.067 8%~0.152 1%, 不同株系间存在一定差异,CC71含量相对较高,而CC70含量较低。2种Bt毒蛋白含量比较分析可知,BtCry3型的毒蛋白表达量非常高,是BtCry1型的10倍。
2.3 转不同外源Bt基因株系对不同害虫的抗虫选择性 2.3.1 转BtCry1Ac+API基因株系对不同害虫的抗虫选择性转BtCry1Ac+API基因741杨对鳞翅目害虫的抗虫效果已有很多报道。转BtCry1Ac+API基因株系pb29对杨扇舟蛾、舞毒蛾(Lymantria dispar)、美国白蛾(Hyphantria cunea)等鳞翅目食叶害虫具有高抗虫性,幼虫死亡率均在80%以上,并且还能抑制存活下来的昆虫幼虫的发育,使其发育速率减缓,不能正常结茧,连续4年饲虫试验未发现其抗虫性有规律下降的趋势。Pb17具有中等抗虫性,对杨扇舟蛾等鳞翅目幼虫致死率50%~80%(Yang et al., 2003;杨敏生等,2005)。
用这2个株系饲养鞘翅目害虫柳兰叶甲幼虫,结果列于图 5。随着饲养时间的延长,幼虫的死亡率增加,饲养到第4天时,幼虫死亡率达到28.4%~42.8%,但转基因株系与未转基因的对照没有明显差异,且各株系幼虫死亡率的变异幅度均较大。表明人工饲养柳兰叶甲幼虫,其敏感性较高,自然死亡率较高,但试验结果未表现出Bt毒蛋白的抗虫效果。

|
图 5 柳蓝叶甲幼虫取食转BtCry1Ac+API基因741杨叶片的死亡率 Figure 5 The mortality rate of P. versicolora larva breeded by the leaf of transgenic poplar clone 741 carrying BtCry1Ac+API gene |
用不同株系转抗虫基因(BtCry3A)741杨叶片饲养鞘翅目害虫柳兰叶甲幼虫,以未转基因741杨作对照,结果如表 1。
|
|
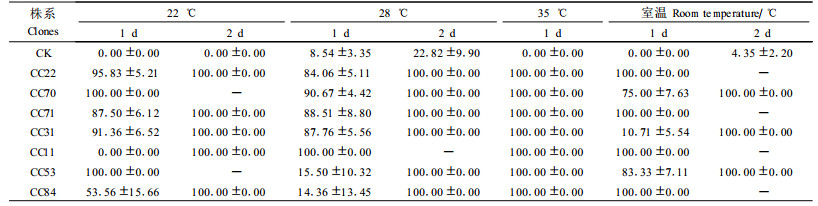
由表 1可以看出,转基因的不同株系对柳蓝叶甲幼虫的致死作用均很强,部分株系饲养的柳蓝叶甲幼虫在第1天的死亡率就为100%,第1天的死亡率未达到100%的第2天均达到100%。所以不同株系间的致死效应均很强,而不同株系对柳蓝叶甲幼虫的死亡率影响不是很大,均在1~2 d之内死亡率达到100%。由此可见,各转基因株系对柳蓝叶甲幼虫均属于高抗性的,死亡率在2 d之内均可达到100%。
为研究Bt毒蛋白在不同温度环境下的表达情况,设置了4个环境温度进行饲虫试验。从表 1结果可以看出,在22℃条件下,幼虫的死亡速率比较缓慢,除了2个株系外,大部分株系需要2 d才能将柳兰叶甲幼虫全部杀死。随着环境温度的提高,到35℃时,所有株系均在1 d内将幼虫杀死,说明在一定温度范围内,高温有利于Bt毒蛋白的表达。
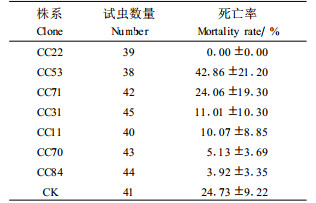
用不同株系的转抗虫基因(BtCry3A)741杨叶片饲养杨扇舟蛾幼虫14 d,以未转基因741杨作对照,观察结果表明:不同株系转抗虫基因(BtCry3A)741杨对杨扇舟蛾幼虫死亡率有不同的影响。从表 2可以看出:用未转基因741杨叶片喂养的幼虫死亡率居中,死亡率24.73%;转抗虫基因741杨CC84,CC70,CC11,CC31饲养幼虫的死亡率比对照741杨低,死亡率为3.92%~11.01%;CC71饲养幼虫的死亡率与对照差异较小,死亡率为24.06%;CC53饲养幼虫的死亡率比普通741杨的要高,死亡率为42.86%;用CC22的叶片饲养杨扇舟蛾幼虫,死亡率为0。由此表明,转抗虫基因(BtCry3A)741杨各株系对鳞翅目害虫杨扇舟蛾没有明显抗性,仅CC53的死亡率略高于未转抗虫基因的741杨。由于限于条件每个重复饲养的昆虫数量少,不同重复之间幼虫死亡率差异较大,经方差分析不同株系之间未达到显著差异。
|
|
自从1981年第1个杀虫晶体蛋白基因被克隆和测序以来,至今已克隆了几百个杀虫晶体蛋白基因。根据杀虫晶体蛋白的大小及基因特点,主要分为Cry和Cyt两大类。目前广泛应用的主要为Cry类杀虫晶体蛋白。Hofte等(1989)根据杀虫蛋白质氨基酸序列的相似性和杀虫活性,将其分成数个类型,分别用罗马数字Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ等来命名。不同类型Cry晶体蛋白具有不同杀虫选择性,Cry为鳞翅目特异性,CryⅡ为鳞翅目和双翅目特异性,CryⅢ为鞘翅目特异性,CryⅣ为双翅目特异性。在每一类型下根据氨基酸序列的同源性,又分为A、B、C等不同的基因型。
许多报道证明转不同类型Bt基因表现出专一抗虫性特征,如转BtCry1的转基因杨树对杨扇舟蛾、舞毒蛾等害虫表达出抗虫效果,而转BtCry3基因杨树对叶甲等鞘翅目害虫具抗虫性,但对其他害虫的试验结果未见报道。为了检测转入植物体中的BtCry1和BtCry3基因对昆虫抗虫性的选择性,本试验用转双抗虫基因(BtCry1Ac+API)741杨饲养柳蓝叶甲幼虫;用转抗虫基因(BtCry3A)741杨饲养鳞翅目昆虫杨扇舟蛾幼虫,结果表明:转双抗虫基因(BtCry1Ac+API)741杨对柳蓝叶甲幼虫的成活没有任何影响,柳蓝叶甲幼虫能完整地完成1个世代,即由幼虫发育为成虫。转抗虫基因(BtCry3A)741杨对杨扇舟蛾幼虫,同一个试验过程中,饲养的杨扇舟蛾幼虫的死亡率与用普通741杨饲养的幼虫死亡率相差不大,其中有些株系比对照的死亡率还要小。而经过数年试验证明的转双抗虫基因741杨对杨扇舟蛾表现高致死性(Yang,2003;杨敏生等,2005)。试验结果进一步证明了bt毒蛋白的专一抗虫性特征,而对非靶标昆虫没有毒杀作用。
转Bt基因对非目标昆虫、土壤微生物、哺乳动物及人类等影响的研究方面,己取得阶段性成果(苏晓华等,2003)。转不同抗虫基因的杨树对昆虫具有一定的选择性,对非靶标昆虫没有毒害作用,这从一方面说明转Bt基因树木具有杀虫谱窄,针对性强的特点,从另一方面也说明其安全性是比较高的,不会对非靶标害虫及其他生物造成危害。鉴于转Bt树木的杀虫特点,建议在生产中应用时,应根据当地虫害特点选择使用不同类型的转基因树木,既在鞘翅目昆虫为害严重的地区栽种转BtCry1基因高抗鞘翅目害虫的树木,而在鳞翅目昆虫为害严重的地区栽种大量转Btcry3基因高抗鳞翅目害虫的树木。为了扩大杀虫谱,提高抗虫效果,将转不同类型Bt基因树木混合栽种,或者将不同类型Bt基因构件在一个表达载体上,通过基因转化,获得转多Bt基因树木,在生产上栽植应用,通过多种抗虫基因抗性互补,可大大提高转基因树木的抗虫能力,这应该是在今后重点研究的方向。
郭同斌, 嵇保中, 诸葛强, 等. 2004. 转Bt基因杨树(NL-80106)对杨小舟蛾抗虫性研究. 南京林业大学学报, 28(6): 5-9. DOI:10.3969/j.issn.1000-2006.2004.06.002 |
李海涛, 王洪成, 刘志洋. 2004. Bt杀虫晶体蛋白的研究概述. 黑龙江农业科学, (5): 37-39. DOI:10.3969/j.issn.1002-2767.2004.05.014 |
苏晓华, 张冰玉, 黄秦军, 等. 2003. 我国林木基因工程研究进展及关键领域. 林业科学, 39(5): 111-118. DOI:10.3321/j.issn:1001-7488.2003.05.017 |
田颖川, 郑钧宝, 虞红梅, 等. 2000. 转双抗虫基因杂种741毛白杨的研究. 植物学报, 42(3): 263-268. |
王关林, 等. 2002. 植物基因工程. 2版. 北京: 科学出版社.
|
王敏杰, 卢孟柱. 2002. 林木基因工程育种现状与发展趋势. 世界林业研究, 15(3): 7-13. DOI:10.3969/j.issn.1001-4241.2002.03.002 |
王学聘, 韩一凡, 田颖川, 等. 1997. 抗虫转基因欧美杨的培育. 林业科学, 33(1): 69-74. DOI:10.3321/j.issn:1001-7488.1997.01.010 |
杨敏生, 高宝嘉, 王进茂, 等. 2005. 转双抗虫基因741杨基本特性分析. 林业科学, 41(1): 91-97. |
张冰玉, 苏晓华, 李义良, 等. 2006. 转抗鞘翅目害虫基因银腺杨的获得及其抗虫性的初步研究. 北京林业大学学报, 28(2): 102-105. DOI:10.3321/j.issn:1000-1522.2006.02.018 |
Balestrazzi A, Allegro G, Confalonieri M. 2006. Genetically modified trees expressing genes for insect pet resistance// Fladung M, Ewald D. Tree transgenesis: Recent developments. Berlin: Springer, 253-274. https://link.springer.com/chapter/10.1007%2F3-540-32199-3_12
|
Hofte H, Whiteley H R. 1989. Insecticidal crystal protein of Bacillus thuringiensis. Microbiolo Rev, 53(2): 242-255. |
Van Frankenhuyzen K, Beardmore T. 2004. Current status and environmental impact of transgenic forest trees. Can J For Res, 34(6): 1163-1180. DOI:10.1139/x04-024 |
Yang M S, Liang H Y, Gao B J, et al. 2003. Insecticidal activity and transgene expression stability of transgenic hybrid poplar clone 741 carrying two insect-resistant genes. Silvae Genetica, 52(6): 197-201. |
Zhang Q, Zhang Z Y, Lin S Z, et al. 2005. Resistance of transgenic hybrid triploids in Populus tomentosa Carr. against 3 species of lepidopterans following two winter dormancies conferred by high level expression of cowpea trypsin inhibitor gene. Silvae Genetica, 54(3): 108-116. |