 2008, Vol. 44
2008, Vol. 44文章信息
- 曾立雄, 王鹏程, 肖文发, 万睿, 黄志霖, 潘磊.
- Zeng Lixiong, Wang Pengcheng, Xiao Wenfa, Wan Rui, Huang Zhilin, Pan Lei
- 三峡库区主要植被生物量与生产力分配特征
- Allocation of Biomass and Productivity of Main Vegetations in Three Gorges Reservoir Region
- 林业科学, 2008, 44(8): 16-22.
- Scientia Silvae Sinicae, 2008, 44(8): 16-22.
-
文章历史
- 收稿日期:2008-01-21
-
作者相关文章



2. 中国林业科学研究院 北京 100091;
3. 湖北省林业科学研究院 武汉 430075
2. Chinese Academy of Forestry Beijing 100091;
3. Forestry Science Institute of Hubei Province Wuhan 430075
森林生物量和生产力是生态系统性质、状态的重要指示特征,也是深入了解森林生态系统变化规律的重要途径(冯宗炜等,1999)。森林生物量和生产力的研究越来越受到研究者的重视(罗天祥等,1999;Keller et al., 2001;Chave et al., 2003;Dewalt et al., 2004)。
林分生物量与生产力的空间结构是指林分不同层次生物量和生产力在空间上的配置状况(Brown et al., 1999;吕晓涛等,2007),它在很大程度上反映了植物对光能、水分和养分的利用效率(彭少麟等,1994),对研究植被碳储量的分布具有重要意义(Falkowski et al., 2000;康冰等,2006)。目前生物量的分配研究较多(樊后保等,2006;康冰等,2006;吕晓涛等,2007),而对各营养器官生物量比例与单株生物量之间以及营养器官生物量比例之间相对生长规律的研究较少。本文研究了整个三峡库区主要植被类型生物量和生产力的分配及各器官的相对生长规律,以进一步了解不同树种在生长过程中对环境资源的利用规律,为研究三峡库区植被生物量、生产力以及碳储量的空间分布格局提供基础数据。
1 研究区概况长江三峡库区(105°49′—111°39′ E,28°28′—31°44′ N)东起湖北宜昌,西至重庆江津,涉及20个县(市、区),总面积5.8万km2。地处亚热带的北缘,具亚热带湿润性气候特征,整个库区四季分明,年降水量1 000~1 250 mm,夏季热而多雨,4—10月降水量占全年降水量的80%以上。区内热量充足,年均气温17~19 ℃,极端最高温可达44 ℃,极端最低气温-2.5~5.6 ℃,全年无霜期300~340 d。相对湿度较大(60%~80%)。受地形的影响,局地气候明显,年均温和年降雨量高于同纬度的中国东部地区。
库区横跨鄂西、渝东山地,地势复杂,海拔跨度大,该区域植物种类繁多,结构复杂。其中针叶林所占森林面积最大,针叶树种主要有马尾松(Pinus massoniana)、柏木(Cupressus funebris)和杉木(Cunninghamia lanceolata);落叶阔叶树以栎属(Quercus)、枫香(Liquidambar formosana)和刺槐(Robinia pseudoacacia)为主;常绿阔叶林中石栎(Lithocarpus glaber)、青冈(Cyclobalanopsis glauca)和栲(Castanopsis fargesii)所占比重较大。
受岩性的影响,库区内有较大面积发育在紫色砂页岩上的紫色土及发育在石灰岩上的各类石灰土(钙质土),石灰岩地区在海拔1 400 m以下主要分布有山地黄壤,为本区地带性土壤,海拔1 500 m以上分布有山地黄棕壤。
2 研究方法 2.1 样地设置肖文发等(2000)和程瑞梅等(1999)曾对库区内植被做过系统整理和分类。三峡库区主要植被类型包括暖温性常绿针叶林(暖性松林、柏木林、杉木林)、典型落叶阔叶林(栎类混交林)、一般落叶阔叶林(其他软阔林)、典型常绿阔叶林(其他硬阔林)、暖温性针阔混交林(马尾松针阔混交林、杉木针阔混交林)、温性常绿针叶林(温性松林)、竹林、灌木林和柑橘(Citrus reticulata)人工林等。
结合1999年三峡库区森林资源2类清查资料,确定本研究选取的10种森林类型(表 1)。在所选择森林类型的主要分布区内按经度、纬度、海拔等因子差异设置临时样地104块,每块样地面积为400~600 m2。应注意尽量选择树木径级分布均匀的乔木林样地,及无人为破坏的灌木和竹林样地。
|
|

在样地范围内进行每木检尺调查,以平均胸径为标准选取1~2株标准木。分别在树干的基部,1.3和3.6 m处锯断,之后每隔2 m锯断,各段称带皮和去皮鲜质量,对于每一段树干和树皮分别取样。分层、分级调查枝叶生物量,从第一活枝起,将树冠等分成上、中、下3层,然后在各层内以枝基径<1 cm、1~2 cm、2~4 cm与>4 cm为标准进行分级称鲜质量和取样。由于本次调查树种结实量小,因而将果实算做枝质量,没有单独采集。地下部分生物量测定按不同方向(上、下、左、右)及不同层次(0~30 cm、30 cm以下)挖掘根系,分细根(d<1.0 cm)、中根(1.0~2.0 cm)、粗根(>2.0 cm)和根桩4类,分层分类称重后,各自取样(樊后保等,2006)。竹子根系生物量没有调查。林下植被生物量是在样地内分别于四角和中央共取5个2 m×2 m的小样方,将小样方内所有林下植被连根全部挖出,称鲜质量并取样(刘国华等,2003)。收集样地四角和中央5个0.5 m×0.5 m小样方的枯落物,称鲜质量并取样。所有样品带回实验室在85 ℃烘箱内烘至恒质量,求出各样品含水率,然后依据鲜质量求出各器官、各部位的干质量。
根据实测数据建立生长预测模型,结合每木检尺计算样地乔木层生物量,根据面积换算林下层生物量和枯落物层现存量。
由于所研究林分主要为中龄林和幼龄林,枯立木和病虫害比较少,所以未考虑枯立木和病虫害对生物量的影响。
2.3 相对生长方程建立选择近年来使用较多的生物量模型对所有树种及营养器官分别进行模拟(王鹏程等,1998)。各树种整株与其组成部分选择相同模式的模型,以树种整体生物量模型的检验精度作为主要选择标准,在树种整体生物量模型检验精度都较高的情况下(P<0.001),则选择各组成部分模型检验精度最高的模型。各树种生物量预测模型见表 2。
|
|
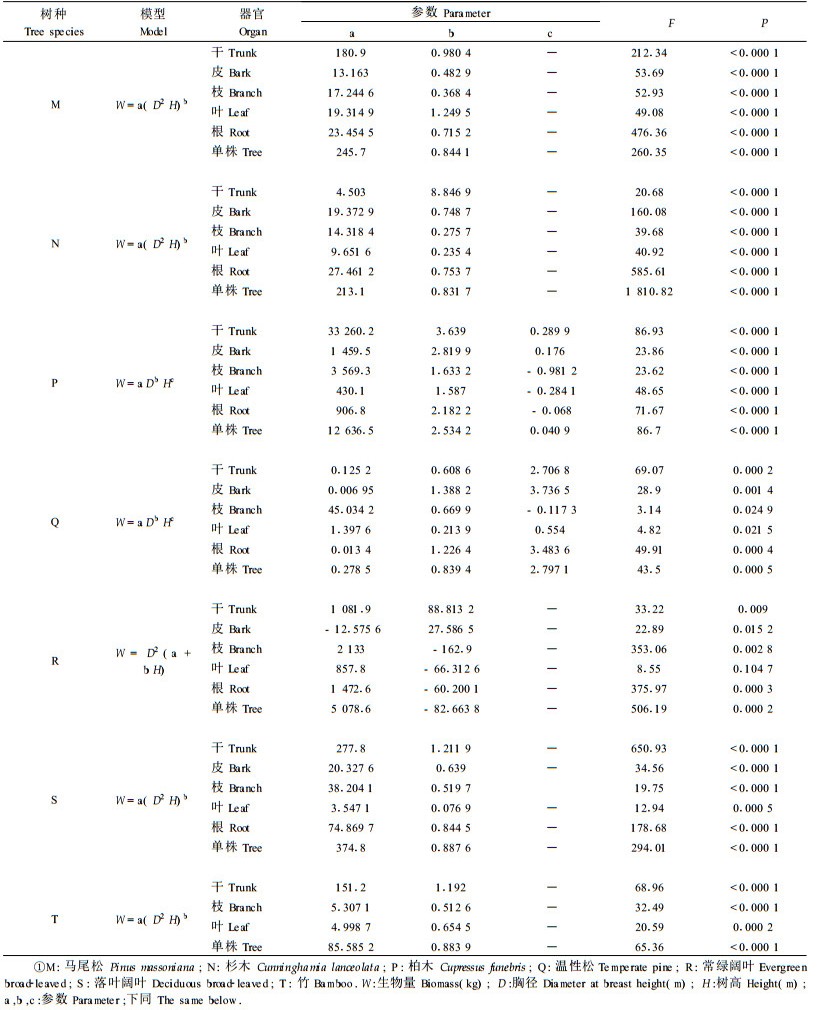
树种都具有自己的生长特性,不同树种其营养器官生物量所占比例的大小顺序也有所差异。如表 3所示,杉木、常绿阔叶和落叶阔叶树种各部分生物量所占比例大小顺序为干>根>枝>皮>叶;马尾松为干>枝>根>皮>叶;柏木为干>枝>根>叶>皮;温性松为干>根>皮>枝>叶;竹子为干>枝>叶。可见,各树种干的生物量都明显高于其他营养器官,除柏木外叶生物量最小,而皮、枝、和根生物量的比例随树种不同其大小顺序会发生改变。
|
|
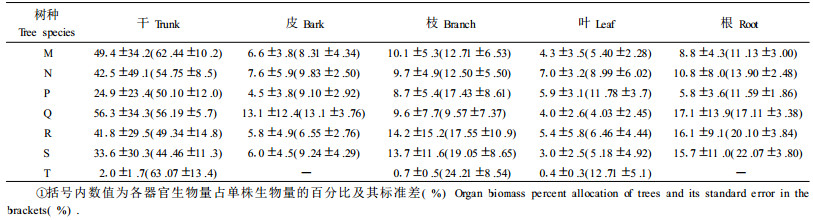
对于同一营养器官,树种不同其生物量所占比例也有较大差别,干生物量中马尾松和竹类所占比例较大,达到62.44%,落叶阔叶树种最低,只有44.46%。其中针叶树种干生物量的比例都比阔叶树种高,而阔叶树种枝生物量的比例则普遍高于针叶树种,这主要是由于针叶树种主干比较明显,而阔叶树种分枝比较多;皮生物量中温性松所占比例最大,其他树种之间差别不大;叶生物量中柏木所占比例明显大于其他树种,杉木其次;根生物量中也是阔叶树种明显高于针叶树种。各针叶树种地上部分生物量占总生物量的比例为82.89%~88.87%,阔叶树种则不到80%。
3.2 营养器官生长规律 3.2.1 营养器官与单株生物量之间的分配规律一般说来,随着树高和胸径的生长,单株林木生物量会增加,林木各器官生物量也会增加(谢宗强等,1999;张光灿等,2004)。但各器官生物量占单株生物量的比例如何变化却没有详细的研究,器官生物量比例的变化可以反映其在林木生长过程中生长速度的快慢,也有助于理解林木单株生物量分配规律的动态变化。
整体来看,随着树木的生长,干和皮的比例会随之增加,枝、叶和根的比例则有所下降。不同树种之间的生长速度存在一定的差别,马尾松、杉木、柏木和温性松等针叶树种各器官比例与单株生物量相关性较高,表现出很好的生长规律,而阔叶树种和竹子各器官比例的生长波动性较大,与单株生物量的增长没有明显的线性关系(表 4)。
|
|
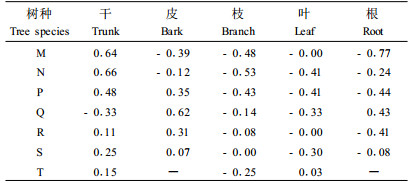
从各个器官来看,马尾松、杉木、柏木和温性松的树干比例与单株生物量的相关性明显高于阔叶树种和竹子。除了温性松,其他树种的干生物量比例都随单株生物量的增加而增加,结合表 3发现温性松树皮生物量所占比例过高,且与单株生物量显著正相关,这可能是造成温性松树干生物量比例随单株生物量增加而减少的原因之一。所有树种的枝生物量比例都与单株生物量成负相关,即随着林木的生长,其枝生物量所占的比例会越来越小,尤其是马尾松、杉木和柏木等针叶树种会出现明显的下降。叶比例的变化规律与枝比较相似,除竹子外都随林木的生长而比例下降。马尾松的根比例与单株生物量显著负相关,除温性松外其余树种的根也都随着林木的生长而比例降低。
3.2.2 各营养器官之间的生长规律各器官生物量比例之间的相互关系能够充分反映林木生长过程中的内部动态。表 5为不同树种各器官生物量比例间相关系数的统计值。可以看出,各树种干生物量的比例与枝叶生物量的比例间存在显著的线性负相关,而枝和叶之间则存在明显的正相关,即当干生物量的比例增加时,枝和叶的比例都会明显下降;而枝生物量的比例增加时,叶的比例会随之提高。其他器官之间虽也具有一定的相关性,但规律不明显。
|
|

从树种来看,柏木、温性松、常绿阔叶林和竹子各器官生物量比例之间存在较高的相关性,单株生物量的空间分布规律较强;而马尾松、杉木和落叶阔叶树种除干、枝和叶之间的相关性较高外,其他器官生物量之间互相影响的作用不是很明显。
3.3 林分生物量的层次结构如表 6所示,马尾松林、杉木林、柏木林、温性松林、针叶混交林、针阔混交林、常绿阔叶林、落叶阔叶林、竹林、灌木林以及经济林的林分生物量分别为123.33,173.68,75.38,191.82,159.27,148.55,273.47,124.44,77.44,18.67与41.01 t·hm-2。但无论是哪种林分类型,其林分内各层次之间的平均生物量始终是乔木层>枯落物层>灌木层;其中,温性松林乔木层生物量占林分生物量的比例高达93.32%,略高于其他林分,而马尾松林、柏木林和常绿阔叶林乔木层生物量的比例相对较少,都不到90%;常绿阔叶林灌木层的生物量达到8.35 t·hm-2,明显高于其他乔木林分,但柏木灌木层生物量占整个林分的5.12%则是所有林分中最高,而杉木林灌木层生物量仅有2.40 t·hm-2,占整个林分的比例也只有1.38%,所有林分中均为最低。灌木林灌木层的生物量为11.16 t·hm-2,高于所有乔木林的灌木层生物量;在枯落物层中,杉木林的枯落物生物量最高,为11.61 t·hm-2,其次为常绿阔叶林和温性松林,柏木林的枯落物生物量最低,只有3.95 t·hm-2,灌木林的枯落物生物量虽然只有7.51 t·hm-2,但其占整个林分的比例达到40.22%,为所有林分中最高,比例最低的是针阔混交林,只有4.25%。
|
|
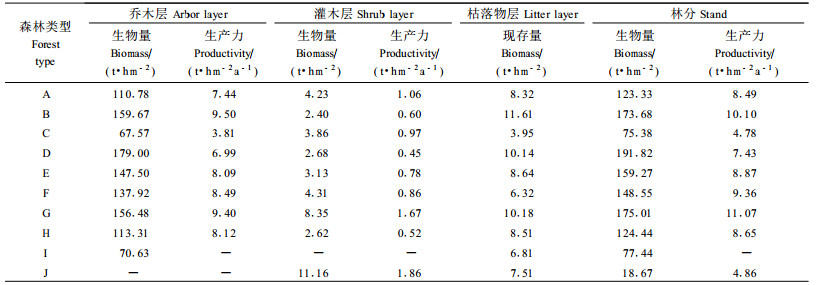
本研究用林木生物量的年均增长量来代替年净生产量,为减小误差,将枯落物归还到乔木层中进行统计。如表 6所示,马尾松林、杉木林、柏木林、温性松林、针叶混交林、针阔混交林、常绿阔叶林、落叶阔叶林以及灌木林林分生产力分别为8.49,10.10,4.78,7.43,8.87,9.36,19.63,8.65与4.86 t·hm-2a-1。除马尾松林、柏木林和常绿阔叶林外,乔木层生产力都达到了林分生产力的90%以上,而各林分灌木层的生产力大部分都低于1.00 t·hm-2a-1,可见乔木层对周边环境中能源的利用量要远多于灌木层。其中,常绿阔叶林乔木层生产力最高,为17.96 t·hm-2a-1,其次为杉木林,柏木林乔木层生产力最低,只有3.81%。
4 结论与讨论1) 所研究树种的单株生物量空间分配中,树干的生物量都明显高于其他营养器官,与樊后保等(2006)、吕晓涛等(2007)的研究结果相同。除柏木外,其他树种都是叶生物量所占比例最少,而皮、枝和根生物量的比例则具有一定的波动性,树种不同其生物量比例的大小顺序也会有所改变,由生物量比例的分析可知,不同生长阶段,树种各器官生物量比例有所差异,由于本研究所选择的树种分布在不同生长阶段,这些可能会对树种各营养器官生物量比例的大小顺序产生一定影响。对于同一营养器官,树种不同其生物量所占比例也有较大差别。其中针叶树种干生物量的比例(50.10%~62.44%)都比阔叶树种(44.46%~49.34%)高,而枝和根的比例则恰好相反,阔叶树种枝和根的比例分别为17.55%~19.05%和20.10%~22.07%。
2) 马尾松、杉木、柏木和温性松等针叶树种各器官生物量的比例与单株生物量的相关性较高,表现出较明显的生长规律。随着单株林木生物量的增加,马尾松、杉木和柏木干和皮生物量的比例会随之增加,枝、叶和根生物量的比例则有所下降;温性松则是皮和根生物量的比例会随之增加,干、枝和叶的比例随之减少。而阔叶树种和竹子各器官生物量比例的生长随机性较大,与树木的生长没有明显的线性关系。在各营养器官生物量比例与其单株生物量相关关系的比较中,整体上关系明显。但其中也有少量不一致的结果,马尾松和杉木皮生物量的比例随其单株生物量的增加而减小,且这2个树种干和皮生物量比例间也是负相关,也与其他树种相反。这种差异是由于这2个树种自身生长特性的不同还是野外数据采集的误差,现在无法判断。另外,随着单株生物量的增加,温性松干生物量的比例在下降,而根的比例则增加,这2点都与其他树种相反。本研究中,温性松包括华山松(Pinus armandii)和巴山松(P.henryi),尽管从植被分类的角度他们都划分为温性松,但2者在生物量的分配方面是否存在差异还没有专门的研究。总之,造成以上差异的原因现在还无法找出准确的答案,还有待在以后的研究中证实,这些将利于更加深入地认识树木在生长过程中的物质分配规律。
3) 本文中所有树种干生物量比例与枝和叶生物量比例间都存在显著的线性负相关,而枝和叶之间则存在明显的正相关,即当干生物量的比例增加时,枝和叶的比例都会明显下降;而枝生物量的比例增加时,叶的比例则会随之提高。其他器官之间虽也具有一定的相关性,但规律都不如上面明显。从树种来看,柏木、温性松、常绿阔叶林和竹子各器官生物量比例之间存在较高的相关性,单株生物量的空间分布规律较强;而马尾松、杉木和落叶阔叶树种除干、枝和叶之间的相关性较高外,其他器官生物量之间互相影响的作用不是很明显。
4) 各林分生物量为75.38~191.82 t·hm-2,乔木层(67.57~179.00 t·hm-2)>枯落物层(3.95~11.61 t·hm-2)>灌木层(2.40~8.35 t·hm-2)。比较发现,枯落物生物量和乔木层生物量低的马尾松林和柏木林,其林下灌木层生物量比较高;而杉木林、温性松林、针叶混交林及落叶阔叶林的枯落物和乔木层数值都比较高的林分,其林下灌木层的生物量比较低;针阔混交林和常绿阔叶林的枯落物生物量和乔木层数值比较高,其林下灌木层生物量也比较高。
5) 各林分生产力为4.78~11.07 t·hm-2a-1,常绿阔叶林最高,柏木林最低,混交林的生产力高于纯林,其中针阔混交林的生产力又高于针叶混交林。
程瑞梅, 肖文发, 李建文, 等. 1999. 三峡库区森林植被分类系统初探. 环境与开发, 14(2): 4-8. |
丁贵杰, 王鹏程. 2001. 马尾松人工林生物量及生产力变化规律研究Ⅱ不同林龄生物量及生产力. 林业科学研究, 15(1): 54-60. DOI:10.3321/j.issn:1001-1498.2001.01.010 |
樊后保, 李燕燕, 苏兵强, 等. 2006. 马尾松-阔叶树混交异龄林生物量与生产力分配格局. 生态学报, 26(8): 2463-2473. DOI:10.3321/j.issn:1000-0933.2006.08.007 |
方精云, 刘国华, 徐嵩龄. 1996. 我国森林植被的生物量和净生产量. 生态学报, 16(5): 497-508. |
冯宗炜, 王效科, 吴刚. 1999. 中国森林生态系统的生物量和生产力. 北京: 科学出版社, 13-14.
|
康冰, 刘世荣, 张广军, 等. 2006. 广西大青山南亚热带马尾松、杉木混交林生态系统碳素积累和分配特征. 生态学报, 26(5): 1320-1329. DOI:10.3321/j.issn:1000-0933.2006.05.004 |
刘国华, 马克明, 傅博杰, 等. 2003. 岷江干旱河谷主要灌丛类型地上生物量研究. 生态学报, 9(23): 1757-1764. |
罗天祥, 李文华, 赵士洞. 1999. 中国油松林生产力分布格局与模拟. 应用生态学报, 10: 257-261. DOI:10.3321/j.issn:1001-9332.1999.03.001 |
吕晓涛, 唐建维, 何有才, 等. 2007. 西双版纳热带季节雨淋的生物量及其分配特征. 植物生态学报, 31(1): 11-22. DOI:10.3321/j.issn:1005-264X.2007.01.003 |
彭少麟, 张祝平. 1994. 鼎湖山地带性植被生物量生产力和光能利用率. 中国科学(B辑), 24(5): 497-502. |
王鹏程, 庄尔奇, 丁贵杰, 等. 1998. 湖北省马尾松人工林结构模型的研究. 华中农业大学学报, 17(1): 77-81. DOI:10.3321/j.issn:1000-2421.1998.01.016 |
肖文发, 李建文, 于长青, 等. 2000. 长江三峡库区陆生动植物生态. 重庆: 西南师范大学出版社, 5-11.
|
谢宗强, 陈伟烈, 路鹏. 1999. 银衫单株生长规律与种群生物量的研究. 植物生态学报, 23(6): 536-543. DOI:10.3321/j.issn:1005-264X.1999.06.007 |
杨昆, 管东生. 2006. 林下植被的生物量分布特征及其作用. 生态学杂志, 25(10): 1252-1256. DOI:10.3321/j.issn:1000-4890.2006.10.019 |
张光灿, 刘霞, 周泽福, 等. 2004. 黄土丘陵区油松水土保持林生长过程与直径结构. 应用生态学报, 18(4): 728-736. DOI:10.3321/j.issn:1001-9332.2004.04.038 |
Brown S L, Schroeder P E S. 1999. Patial patterns of aboveground production and mortality of woody biomass for eastern U.S. forests. Ecological Applications, 9(3): 968-980. |
Chave J, Condit R, Lao S, et al. 2003. Spatial and temporal variation of biomass in a tropical forest: results from a large census plot in Panama. Journal of Ecology, 91: 240-252. DOI:10.1046/j.1365-2745.2003.00757.x |
Dewalt S J, Chave J. 2004. Structure and biomass of four lowland neotropical forests. Biotropica, 36: 7-9. |
Falkowski P, Scholes R J, Boyle E, et al. 2000. The global carbon cycle: a test of our knowledge of earth as a system. Science, 290: 291-296. DOI:10.1126/science.290.5490.291 |
Keller M, Palace M, Hurtt G. 2001. Biomass estimation in the Tapajos National forest, Brazil examination of sampling and allometric uncertainties. Forest Ecology and Management, (154): 371-382. |