 2008, Vol. 44
2008, Vol. 44文章信息
- 张春雨, 赵秀海, 夏富才.
- Zhang Chunyu, Zhao Xiuhai, Xia Fucai
- 长白山次生林树种空间分布及环境解释
- Spatial Distribution of Tree Species and Environmental Interpretations of Secondary Forest in Changbai Mountains
- 林业科学, 2008, 44(8): 1-8.
- Scientia Silvae Sinicae, 2008, 44(8): 1-8.
-
文章历史
- 收稿日期:2007-07-04
-
作者相关文章



2. 北华大学林学院 吉林 132013
2. Forestry College of Beihua University Jilin 132013
长白山覆盖着大面积未受干扰的温带原始森林,它为虎、熊、豹、狼、野猪等野生动物提供了重要栖息地。原始阔叶红松(Pinus koraiensis)林林龄超过200年,是我国过去几十年间的重要木材产地之一。但20世纪初,大规模工业采伐使原始阔叶红松林遭到了严重破坏,并逐渐为次生林或人工林所代替(Chen et al., 1994)。
虽然对次生林的保护、经营和恢复已经迫在眉睫,但对次生林种群和群落的空间分布规律了解却十分有限。以往研究显示,绝大多数树种在空间上并不是均匀或随机分布,而呈聚集性分布(孙伟中等,1997),这说明某些过程或机制支配着森林空间结构的形成。环境决定论者认为环境变化是影响物种格局的主要根源(环境控制模型),而生物决定论者则认为生物之间的互作是构造群落结构分异的动力(生物控制模型)(May,1984)。通常来说,有2种机制可以制约植物空间结构的形成:1)植物种群的空间自相关,即同种植物种群内不同个体之间的相互吸引或排斥关系。植物的生长、死亡、种子传播以及竞争动态等生物学过程都会影响植物在群落水平上的空间分布状态。2)外在因素作用。当植物种群的环境或生物学控制因素以及历史动态(人类活动、干扰或历史事件)等外在因素存在空间结构性时,会导致植物种群形成一定的空间结构(Legendre et al., 1998),这个过程也被称为诱导空间依赖性(Dray et al., 2006)。Whittaker(1956)与Bray等(1957)认为可以利用环境控制模型对当前的植物分布进行较好地解释。通常群落结构的空间异质性主要来源于环境变量的物理驱动力以及群落结构对环境的空间依赖性,并且空间结构性在生态系统中起了一个功能性作用(Boarcard et al., 2002)。
目前生物种群的空间结构特征、种群空间分布与环境之间的关系、环境变量对种群分布影响等问题已经被生态学家所关注(Brown et al., 1995;1996)。本试验检验了8个环境变量(土壤全氮、土壤全磷、土壤全钾、土壤pH值、土壤水分、土壤有机质、叶面积指数、光量子通量密度)以及12个树种的空间格局(空间自相关性),探讨了环境因子与植物空间分布之间的关系。并试图回答植物种群的空间分布格局是如何形成的,环境因子对植物的空间分布有何影响等问题。
1 研究区概况研究区位于吉林省白河林业局光明林场(128°07′49″ E,42°19′10″ N),海拔899 m。属于受季风影响的温带大陆性山地气候,年均气温3.3 ℃,最热月8月份平均气温20.5 ℃,最冷月1月份平均气温-16.5 ℃,极端最高气温32.3 ℃,极端最低气温-37.6 ℃。年降水量600~900 mm。土壤为山地暗棕色森林土,土层厚度20~100 cm。该区原始植被为阔叶红松林,经采伐破坏后形成针阔混交林,林龄50~70年,乔木种包括红松、蒙古栎(Quercus mongolica)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、色木槭(Acer mono)、春榆(Ulmus japonica)、糠椴(Tilia mandshurica)、簇毛槭(Acer barbinerve)、假色槭(A. pseudo-sieboldianum)、青楷槭(A. egmentosum)、白牛槭(A. mandshuricum)、花楷槭(A. ukurunduense)、怀槐(Maackia amurensis)、拧筋槭(Acer triflorum)、黄菠萝(Phellodendron amurense)、枫桦(Betula costata)、大青杨(Populus ussuriensis)等。
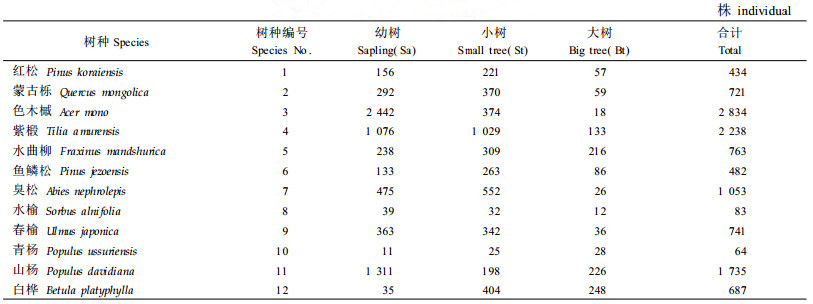
2 研究方法 2.1 数据收集2005年7月建立1个260 m×200 m的固定样地,将其进一步划分为130个20 m×20 m的连续样方。记录样地内所有DBH≥1 cm的乔木个体种名、胸径、树高、冠幅(东西冠幅长、南北冠幅长)、枝下高及其位置坐标。假设树木胸径与生长年龄相对应(Sterner et al., 1986),根据胸径划分幼树(1 cm≤DBH<5 cm)、小树(5 cm≤DBH<20 cm)及大树(DBH≥20 cm)3个生长阶段。本研究选择了12个主要树种,树种概况见表 1。
|
|
在每个样方土壤表层(0~20 cm)各取2个500 g的土壤样品,带回实验室进行分析。室内土壤样品分析方法依据《土壤农业化学分析方法》(中国土壤学会,1999)的操作步骤进行:用开氏消煮法测定土壤中全氮含量,碳酸钠熔融法测定土壤中全磷含量,氢氧化钠熔融法测定土壤中全钾含量,高温外热重铬酸钾氧化-密度法测定土壤中有机质含量,用pH计测定土壤pH值。计算2个样品的平均值作为每个样方的值。用手持土壤水分测定仪(HH2 DelLa-T Devices Moisture Meter,UK.)测定每个样方表层土壤水分含量。为了评价林分光照状况,在2005年8月的无云晴天,使用冠层分析仪(WinSCANOPY,Canada)在每个样方中距离地面1 m处拍摄2张半球状照片(hemispherical photographs),用对应的WinSCANOPY和XLScanopy软件处理照片,计算林下包括直射光和散射光在内的总光量子密度(PPFD),并依据Bonhomme等(1972)的方法测算冠层结构指标叶面积指数(LAI)。LAI值小,光照较强;LAI值大,光照较弱(韩有志等,2004)。由于样地地势平坦,坡度小于5°,地形变化较小,故本文未考虑地形对植被分布的影响。
2.2 数据分析 2.2.1 空间格局分析局部Moran's I统计量是一种常用的检验变量局部空间相关性的统计量。空间自相关系数Moran's I可用如下公式进行计算:
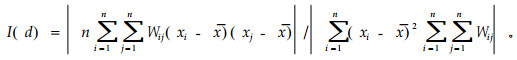
|
在环境变量空间自相关分析中,xi和xj分别为环境因子x在配对空间单元i和j上的观测值,x为环境因子在n个空间单元上含量的平均值。在植物种群空间自相关分析中,xi和xj分别为种群x在配对空间单元i和j上的株数,x为种群在n个空间单元上的株数平均值。Wij为根据空间单元i和j的相对位置计算的二元相邻权重。当空间单元i和j位于相同的行或列,并且它们之间的距离等于d,即空间单元i和j相连接时,Wij=1;否则,Wij=0。本文将样点间最大空间距离(300 m)分为10段,在10个滞后距离级上计算8个环境因子以及12个树种的空间自相关系数Moran's I(d=1,2,…,10),并应用置换(Permutation)检验计算Moran's I的显著性。空间自相关系数Moran's I计算及显著性检验通过R软件的sp.correlogram函数实现。
Moran's I值通常介于1与-1之间,当Moran's I值为0时,表示环境因子或植物种群不存在空间自相关,当具有较高的空间自相关时,其值接近于1或-1。正空间自相关代表相邻空间具有相似的特征或属性,负空间自相关则表示相邻空间具有不同的特征或属性,无空间自相关则表示不存在支配格局形成的特定系统结构(即格局趋向于随机性)。Moran's I值从1(正空间自相关)向-1(负空间自相关)变化,反映了由空间相邻相似的正相关向空间相邻相异的负相关过渡。空间异质性可以导致变量在某一区域上呈正空间自相关,而在另一区域上呈负空间自相关。显著的空间自相关暗示着一个非随机性空间分布格局,即变量存在空间格局。显著正空间自相关代表着聚集性分布,显著负空间自相关代表着均匀性分布格局。有关Moran's I的计算方法及理论解释见参考文献(Sokal et al., 1978a;1978b)。
2.2.2 植被变异分割变异分割(variation partition)是一种用2个或多个解释变量来定量分割反应变量的方法。对于简单变量,变异分割可通过局部线性回归(partial linear regression)实现,对于多变量数据,可应用CCA或RDA方法分割反应变量的变异。当用环境矩阵X和描述样点空间关系的矩阵W来分割物种组成或种群丰富度矩阵Y时,Y的空间变异可被分割为4部分:1)环境因子独立作用部分;2)空间因子和环境因子交互作用部分;3)空间因子独立作用部分;4)空间因子和环境因子不能解释的部分(Legendre et al., 1998)。为了评价空间结构的重要性,Borcard等(1992;1994)早在1992年就将取样点地理坐标多项式(即趋势面)作为空间变量引入统计模型,该方法可以描述大尺度的地形变化,但无法描述小尺度的局部地理变异。但一种新的多变量空间格局分析方法——相邻矩阵主坐标(PCNM)已经被广泛使用,这种方法克服了趋势面无法充分表达小生境特征的缺点(Borcard et al., 2002)。
本文将平方根转化后的种群丰富度数据(定义为每个20 m×20 m亚样方中的株数)定义为反应矩阵Y,由全氮、全磷、全钾、有机质、土壤pH值、土壤水分、叶面积指数和光量子密度这8个变量构成环境矩阵X,相邻矩阵主坐标(PCNM)作为空间矩阵W。应用向前选择法(forward selection)评价解释变量(环境因子和空间变量)与反应变量(种群丰富度格局)间的关系,并通过999次随机置换(permutations)删除冗余变量,选择最佳环境变量及空间变量组合。使用选定的环境变量和空间变量,通过RDA方法来分割反应变量的空间变异。利用调整决定系数R2来估计不同解释部分的贡献率(Peres-Neto et al., 2006)。相邻矩阵主坐标利用PCNM.R函数计算(Dray et al., 2006),用R软件的“vegan"包进行变异分割计算。
3 结果与分析 3.1 环境变量空间自相关分析环境变量空间自相关分析(图 1)表明,全氮(d=1,2,3)、全磷(d=1,2)、全钾(d=1)、土壤pH值(d=1,2)、土壤水分(d=1)、叶面积指数(d=1,4)6个环境因子具有显著的空间自相关结构,在空间上呈显著的聚集性分布。当d为1~5时,滞后距离越小,Moran's I值越高,样点间的空间正相关也越显著。土壤有机质不存在显著的空间自相关结构,在空间上呈随机分布。林地表面的光量子通量密度也未表现出明显的空间结构,这可能与林分处于中龄林阶段,上方林冠尚未郁闭有关。

|
图 1 环境因子空间自相关分析 Figure 1 Spatial autocorrelation analysis of environmental variables ***:P<0.001;**:P<0.01;*:P<0.05;下同The same below. |
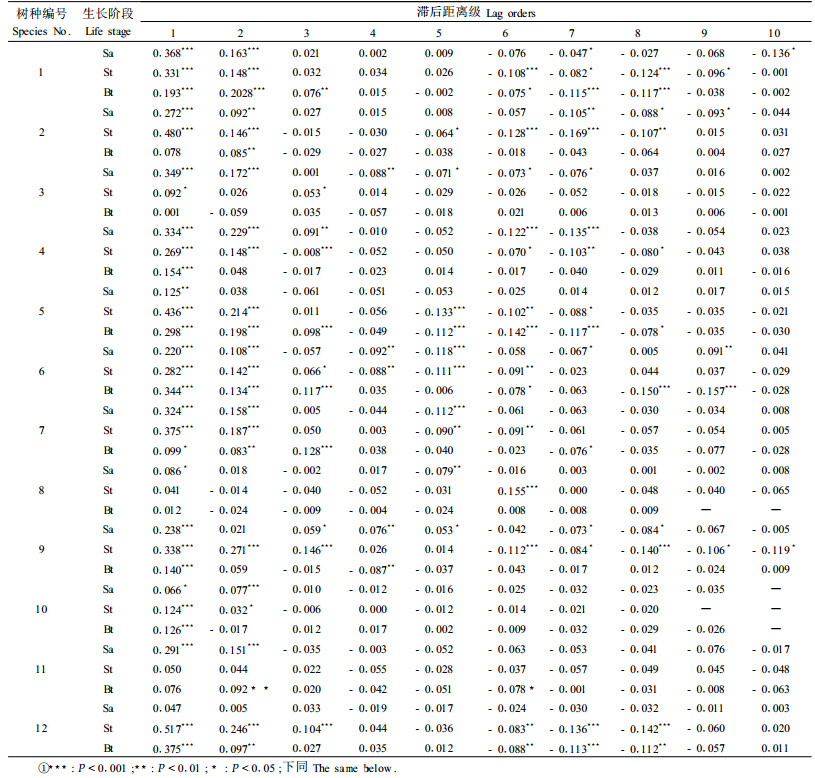
12个树种不同生长阶段(幼树、小树和大树)的空间自相关分析如表 2所示,结果表明白桦幼树、色木槭大树、山杨小树及水榆大树空间自相关不显著(P>0.05),未形成明显的空间分布格局,而其他树种或生长阶段在小距离尺度上(主要表现在d=1,2距离级)均表现出显著的空间正自相关性,说明绝大多数树种呈聚集性空间分布。这与以往针阔混交林树种空间格局研究的结论基本一致(孙伟中等,1997)。树木种群的空间自相关主要来源于植物的生长、补充、死亡、种子传播以及竞争动态等生物学过程,而生物学过程也是植物种群形成空间聚集性结构的一个重要机制。
|
|
由决定系数R2及显著性概率p(通过999次permutation计算)可知,土壤水分含量和土壤pH值是影响树种分布的主要环境因子。土壤水分含量与蒙古栎、紫椴、水曲柳、臭松、山杨和白桦分布显著相关,土壤pH值则与蒙古栎、紫椴、水曲柳、水榆和鱼鳞松分布显著相关。土壤养分(包括土壤全氮、全磷、全钾)则与蒙古栎、色木槭、紫椴、水曲柳、鱼鳞松、臭松、春榆及白桦分布显著相关,而土壤有机质、叶面积指数及光量子通量密度与12个树种的空间分布均无显著关系(P>0.05),故而在变异分割分析中剔除这3个环境因子(表 3)。张元明等也得出了相似结论,认为土壤含水量、土壤pH值是影响塔里木河下游植物群落分布的主要环境因子(张元明等,2004)。植被格局与土壤营养条件并未形成很好的对应关系,这可能与频繁的干扰导致地表不稳定,阻碍了土壤条件对植被的影响有关,干扰成为比土壤营养更为重要的植被格局形成原因(Sakai et al., 1993;杨永川等,2006)。
|
|
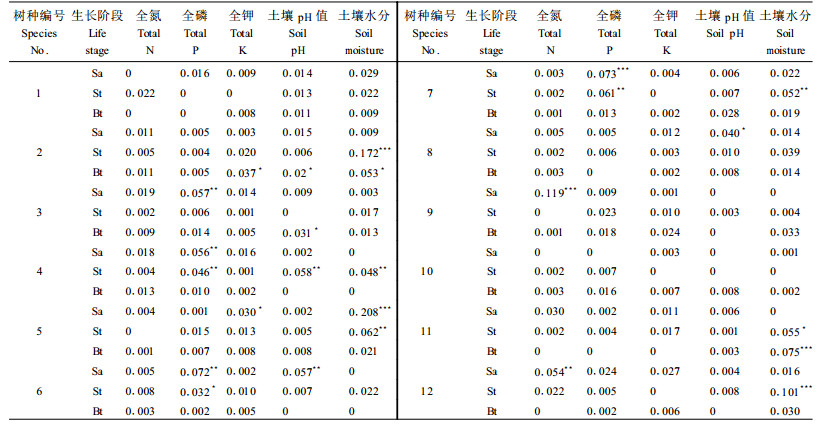
为了进一步评价环境因子对植物种群空间分布的影响,利用解释变量(环境因子及空间变量)对反应变量(树木种群空间分布)进行定量分割。结果表明,红松和青杨的空间分布不受环境因子影响(P>0.05)。环境变量对幼树和小树的空间分布具有较好的解释能力,而对多数树种的大树的解释能力很差。环境因子对蒙古栎、色木槭、山杨大树空间变异的解释能力分别为8.7%,1.7%和6.8%,其他9个树种大树的空间分布与8个环境因子之间均无显著关系(P>0.05)(表 3,4)。环境因子对水曲柳、鱼鳞松和春榆幼树,蒙古栎和紫椴小树空间分布的解释能力均高于10%。水曲柳表现尤为显著,土壤水分和全钾共同解释了水曲柳幼树22.6%的空间变异,其中土壤水分是控制水曲柳幼树分布的主要环境因子(R2=0.208,P<0.001)(表 3,4)。
|
|
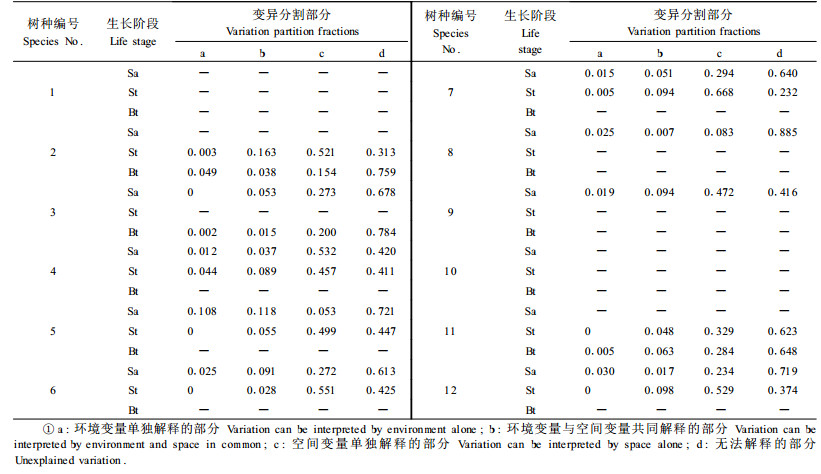
表 3与表 4说明环境因子主要作用于幼树和小树,对大树影响较小。当种群处于不同的发育阶段时,影响其空间分布的外在因素也不尽相同。树木种群在幼树和小树阶段,其分布格局主要受环境因子制约,随着种群发育,植物的生长、死亡及由环境条件引起的种内种间竞争等生物学过程逐渐成为影响大树分布的主导因子。以往研究认为在干旱季节,高光照、高温、低土壤养分和低土壤湿度等微生境特征是树木幼苗、幼树建立和存活的主要限制因子(Uhl et al., 1988;Nepstad,1989;Gerhardt,1993;Hooper et al., 2002),并且对干旱条件(土壤湿度)的敏感性是制约幼苗、幼树空间分布的重要机制(Engelbrecht et al., 2007)。
4 结论与讨论水曲柳幼树的空间分布与土壤水分含量显著相关,在已知的27.9%变异中,仅有5.3%来源于纯粹的空间结构,其他22.6%均来自环境机制。在相关的环境因子中,全钾的局部回归决定系数R2=0.03(P<0.05),土壤水分含量的局部回归决定系数R2=0.208(P<0.001),因此环境因子对水曲柳幼树空间分布的影响主要来源于土壤水分。韩有志等(2004)认为叶面积指数指示的林分光照环境在水曲柳更新中起了关键性作用,林分光照的空间异质性对更新格局起决定性作用。张国仓等(1991)研究表明表层土壤水分状况是影响水曲柳种群天然发生的主要因子,林地上方光照不直接限制水曲柳种群的发生数量,但它可以通过影响种子的萌动速度来影响种群发生进程和数量。本文分析认为土壤水分是决定水曲柳幼树和小树空间分布的主要环境因子,叶面积指数代表的光照条件对水曲柳空间格局没有显著影响(表 3)。
环境因子对红松和青杨的空间分布影响不显著(P>0.05)。这可能与红松分布受动物和人类活动干扰较强烈有关:星鸭(Nucifraga caryocatactes)和松鼠(Sciurus vulgaris)对红松种子的贮藏行为强烈地影响着红松的早期更新格局(Hayashida,1989;Miyaki,1987;鲁长虎等,2001;刘足根等,2005);人类活动,特别是松果采摘造成红松种源缺乏,破坏了动物食物链以及红松种子的传播过程,进而影响到红松种群的空间分布及更新格局(刘足根等,2005)。研究样地中共有64株青杨,其中幼树、小树、大树密度分别为2.1、4.8和5.4株·hm-2(表 1),青杨的分布密度较低,这可能是导致其与环境因子间统计关系不显著的主要原因。
在已知种群空间变异中,环境因子(纯粹的环境因子解释部分以及环境因子和空间变量交互作用解释部分)解释能力较低,而纯粹的空间变量(即未与环境变量共有的空间变量)对种群空间变异具有较高的解释能力,说明一些未知的独立于环境因子的空间过程在树木种群空间结构的形成过程中起了重要作用。这部分空间结构被认为来自于生物性机制或历史事件(Borcard et al., 1992;Legendre et al., 1998)。研究地区林分起源于原始阔叶红松林采伐迹地,且与周围未受干扰或干扰较小的林分相连,种源丰富,因此包含了原始林分的绝大多数物种,但由于林龄较短,林分分布与环境尚未形成良好的适应关系,对环境因子的空间结构也尚未形成诱导空间依赖性(Dray et al., 2006)。空间因子和环境因子未能解释的部分主要反映了群落自身互作及人类活动(采伐或清林等经营行为)对植物种群格局的影响,以及一些未涉及的环境因子、干扰事件或随机因子的影响(Borcard et al., 1992)。
韩有志, 王政权, 谷加存. 2004. 林分光照空间异质性对水曲柳更新的影响. 植物生态学报, 28(4): 468-475. DOI:10.3321/j.issn:1005-264X.2004.04.004 |
刘足根, 姬兰柱, 朱教君. 2005. 松果采摘对种子库及动物影响的探讨. 中国科学院研究生院学报, 22(5): 596-603. DOI:10.3969/j.issn.1002-1175.2005.05.010 |
鲁长虎, 刘伯文, 吴建平. 2001. 阔叶红松林中星鸦和松鼠对红松种子的捕食和传播. 东北林学院学报, 29(5): 96-98. |
孙伟中, 赵士洞. 1997. 长白山北坡椴树阔叶红松林群落主要树种分布格局的研究. 应用生态学报, 8(2): 119-122. DOI:10.3321/j.issn:1001-9332.1997.02.002 |
杨永川, 达良俊. 2006. 丘陵地区地形梯度上植被格局的分异研究概述. 植物生态学报, 30(3): 504-513. DOI:10.3321/j.issn:1005-264X.2006.03.018 |
张国仓, 王义弘, 张雪梅. 1991. 水曲柳实生种群发生的初步研究. 东北林学院学报, 19(5): 26-31. |
张元明, 陈亚宁, 张小雷. 2004. 塔里木河下游植物群落分布格局及其环境解释. 地理学报, 59(6): 903-910. DOI:10.3321/j.issn:0375-5444.2004.06.013 |
中国土壤学会. 1999. 土壤农业化学分析方法. 北京: 中国农业科技出版社.
|
Bonhomme R, Chartler P. 1972. The interpretation and automatic measurement of he mispherical photographs to obtain sunlit folige area and gap frequency. Israel Journal of Agricultural Research, 22: 53-61. |
Borcard D, Legendre P. 1994. Environmental control and spatial structure in ecological communities:an example using orbatid mites (Acari, Orbatei). Environmental and Ecological Statistics, 1: 37-53. DOI:10.1007/BF00714196 |
Borcard D, Legendre P. 2002. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecological Modelling, 153: 51-68. DOI:10.1016/S0304-3800(01)00501-4 |
Borcard D, Legendre P, Drapeau P. 1992. Partialling out the spatial component of ecological variation. Ecology, 73: 1045-1055. DOI:10.2307/1940179 |
Bray J R, Curtis J T. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 27: 325-349. DOI:10.2307/1942268 |
Brown J H, Mehlman D W, Stevens G C. 1995. Spatial variation in abundance. Ecology, 76: 2029-2043. |
Brown J H, Stevens G C, Kaufman D M. 1996. The geographic range:size, shape, boundaries and internal structure. Annual Review of Ecology and Systematics, 27: 597-623. DOI:10.1146/annurev.ecolsys.27.1.597 |
Dray S, Legendre P, Peres-Neto P R. 2006. Spatial modelling:a comprehensive framework for principal coordinate analysis of neighbour matrices (PCNM). Ecological Modelling, 196: 483-493. DOI:10.1016/j.ecolmodel.2006.02.015 |
Engelbrecht B M J, Comita L S, Condit R, et al. 2007. Drought sensitivity shapes species distribution patterns in tropical forests. Nature, 447: 80-83. DOI:10.1038/nature05747 |
Gerhardt K. 1993. Tree seedling development in tropical dry abandoned pasture and secondary forest in Costa Rica. Journal of Vegetation Science, 4: 95-102. DOI:10.2307/3235736 |
Hayashida M. 1989. Seed dispersal by red squirrels and subsequent establishment of Korean pine. Forest Ecology and Management, 28: 115-129. DOI:10.1016/0378-1127(89)90064-9 |
Hooper E, Condit R, Legendre P. 2002. Responses of 20 native tree species to reforestation strategies for abandoned farmland in panama. Ecological Applications, 12(6): 1626-1641. DOI:10.1890/1051-0761(2002)012[1626:RONTST]2.0.CO;2 |
Legendre P, Legendre L. 1998. Numerical ecology. 2nd English ed. Amsterdam: Elsevier Science BV.
|
May R M. 1984. An overview: real and apparent patterns in community structure//Strong D R. Ecological communities: conceptual issues and the evidence. Princeton: Princeton University Press, 3-16.
|
Miyaki M. 1987. Ecology Research. , 147-157.
|
Nepstad D C. 1989. Forest regrowth in abandoned pastures of eastern Amazonia: l imitations to tree seedling survival and growth. Dissertation. New Haven: Yale University Press.
|
Peres-Neto P R, Legendre P, Dray S, et al. 2006. Variation partitioning of species data matrices:estimation and comparison of fractions. Ecology, 87: 2614-2625. DOI:10.1890/0012-9658(2006)87[2614:VPOSDM]2.0.CO;2 |
Sakai A, Ohsawa M. 1993. Vegetation pattern and microtopography on a landslide scar of Mt Kiyosumi, Central Japan. Ecological Research, 8: 47-56. DOI:10.1007/BF02348606 |
Sokal R R, Oden N L. 1978a. Spatial autocorrelation in biology 1. Methodology. Biological Journal of the Linnean Society, 10: 199-228. DOI:10.1111/bij.1978.10.issue-2 |
Sokal R R, Oden N L. 1978b. Spatial autocorrelation in biology 2. Some biologica l implications and four applications of evolutionary and ecological interest. Biological Journal of the Linnean Society, 10: 229-249. DOI:10.1111/bij.1978.10.issue-2 |
Sterner R W, Ribic C A, Schatz G E. 1986. Testing for life historical changes in spatial patterns of four tropical tree species. Journal of Ecology, 74: 621-633. DOI:10.2307/2260386 |
Uhl C, Buschbacher R, Serrao E A S. 1988. Abandoned pastures in eastern Amazonia. Ⅰ. Patterns of plant succession. Journal of Ecology, 76: 663-681. DOI:10.2307/2260566 |
Whittaker R H. 1956. Vegetation of the Great Smoky Mountains. Ecological Monographs, 26: 1-80. DOI:10.2307/1943577 |