 2008, Vol. 44
2008, Vol. 44文章信息
- 李昌晓, 钟章成.
- Li Changxiao, Zhong Zhangcheng.
- 模拟三峡库区消落带土壤水分变化条件下落羽杉幼苗实生土壤营养元素含量的变化
- Nutrient Dynamics in Soils of Taxodium distichum Seedlings under Simulated Soil Water Changes in the Hydro-Fluctuation Belt of the Three Gorges Reservoir Region
- 林业科学, 2008, 44(7): 124-129.
- Scientia Silvae Sinicae, 2008, 44(7): 124-129.
-
文章历史
- 收稿日期:2007-08-08
-
作者相关文章



2. 三峡库区森林生态保护与恢复重庆市市级重点实验室 重庆 400036
2. Chongqing Key Laboratory for the Protection and Restoration of Forest Ecology of the Three Gorges Reservoir Region Chongqing 400036
三峡库区消落带土壤淹水的周期性常年变化,势必影响到土壤营养元素含量、土壤肥力、土壤理化性质等土壤特性的变化(Ponnamperuma,1972;李其林等,2004;杨刚,2004),但土壤营养元素含量的影响是导致土壤其他特性发生相应变化的根本原因之一(金为民,2003;岳庆玲等,2007)。土壤淹水变化引起的土壤营养元素含量变化,将直接影响到植物的生长发育。相反,植物的适应性生长发育也会反过来影响到土壤营养元素含量乃至土壤特性的变化。在三峡库区消落带特定环境条件下,消落带土壤营养元素含量变化不仅有土壤水分的变化引起,也有植物的适应性生长发育引起,从而增加了土壤特性变化的诱因(杨小波等,2000;万福绪等,2006;何斌等,2007)。认识三峡库区消落带植物实生土壤理化性质变化规律,阐明在适生植物生长条件下实生土壤的营养元素动态变化机制,对于消落带植被恢复与水土保持建设具有较大参考应用价值。有关针对三峡库区消落带植物实生土壤营养元素含量变化的还少有报道。
落羽杉(Taxodium distichum)具有很强的耐水湿性能(Randall et al., 2005),在我国长江流域多有引种(汪贵斌等,2002),目前已被建议列为三峡库区消落带的防护林体系建设树种之一(李昌晓等,2005)。因此,本研究通过模拟三峡库区消落带土壤水分变化,旨在认识落羽杉实生土壤营养元素含量的变化机制,为当地植被恢复建设提供技术和理论支持。
1 材料与方法 1.1 研究材料本试验选择落羽杉当年实生幼苗的生长土壤作为研究对象。将生长基本一致的120株幼苗带土盆栽(供试土壤为紫色土,基本理化性质与营养元素含量见表 1),每盆1株。盆中央内径为13 cm,盆内土层厚度12 cm。将所有盆栽试验用苗置于原西南师范大学生态试验园地中(海拔249 m)进行相同土壤基质、光照和水分管理适应,1个月后搭建透明塑料遮雨棚,正式开展水分处理试验。
|
|

将试验用土随机分成4组,每组30盆,包括对照组CK、轻度干旱组T1、水分饱和组T2和水淹组T3。CK即为常规生长组,土壤含水量为田间持水量的60%~63%(土壤含水量采用称重法测定),落羽杉幼苗在晴天无萎蔫现象。T1组为轻度水分胁迫,土壤含水量为田间持水量的47%~50%,植株嫩叶在晴天13:00左右出现萎蔫,17:00左右恢复正常(胡新生等,1998;胡哲森等,2000)。T2组为表面一直处于潮湿状态的水饱和土壤。T3组在本试验中为苗木根部土壤全部淹没,淹水超过土壤表面1 cm。水淹处理时,将苗盆放入直径为68 cm、高22 cm的大型塑料盆内,向盆内注水,直到盆内水面超过土壤表面1 cm为止(Bragina et al., 2001)。从试验处理之日,每间隔5 d为1个处理期,对各项指标连续进行5次测定,每处理每次测定5个重复。
1.3 土壤样品采集及土壤营养元素含量测定取样时先把植株的地上部分割下,将盆钵中的土壤倒出,再将土壤中的所有根系完全取出,只剩下实生土壤。然后将每盆土壤单独充分混合均匀,在室内风干后研磨、过筛、贮存,以供测试。
土壤有机质含量测定采用油浴加热-重铬酸钾容量法;土壤pH测定采用电位计法;土壤全N测定采用全自动凯氏定氮仪(瑞士Büchi公司生产的Distillation Unit B-324)、水解N含量采用水解扩散法;土壤全P与速效P含量测定采用钼锑抗比色法;土壤全K与速效K测定采用火焰光度法。以上土壤理化性质的测定均采用《土壤农业化学常规分析方法》(中国土壤学会农业化学专业委员会,1983)中的有关方法。
1.4 统计分析根据测定生理指标,将水分处理作为独立因数,用一元方差分析(One-way ANOVA)揭示水分变化对落羽杉幼苗实生土壤营养元素含量的影响(GLM程序,SPSS 10.0版)。用Duncan检验法进行多重比较,检验每个指标在处理间(α=0.05)的差异显著性(杜荣骞,2003)。
2 试验结果 2.1 土壤有机质与pH值的变化不同水分处理对落羽杉幼苗实生土壤有机质含量与pH值有极显著影响(表 2)。随着水分处理时间的不断延长,T2组土壤有机质含量与CK组一样,表现出先升后降的变化趋势,而与T3组先降后升的变化趋势形成鲜明对比(图 1)。与之不同的是,T1组土壤有机质含量则表现出升降交替的变化趋势。在整个试验期间,T1、T2和T3组土壤有机质平均含量相互之间均无显著差异,但依次极显著地高于CK组5.8%、9.1%和9.9%(T1与CK组:P < 0.05;T2与CK组:P < 0.05;T3与CK组:P < 0.05)。
|
|


|
图 1 不同水分条件下落羽杉幼苗实生土壤有机质含量与pH值的变化(±标准误) Figure 1 The change of percentage of organic matter and pH value in soils of T. distichum seedlings under different water treatment (±SE) |
土壤pH值在T2组一直连续上升,第5次测量值比第1次高出7.6%,这与CK组pH值升降交替变化趋势明显有所不同。而T3组先升后降的变化趋势与T1组先降后升的变化趋势恰好相反。T2组在整个试验期间的pH平均值与CK组相比并未出现显著差异,但T1与T3组却显著低于CK组,分别降低2.9%和1.7%(T1与CK组:P < 0.05;T2与CK组:P < 0.05;T3与CK组:P < 0.05)。
2.2 土壤全N、全P和全K含量的变化土壤全氮(N)含量在不同水分梯度下有极显著差异存在,与各组全P、全K含量没有显著差异存在迥然不同(表 2)。各处理组在试验期间的全N、全P、全K含量变化趋势各异,基本上是以升降交替形式出现波动性变化。随着处理时间延长,T1组的土壤全N、全P、全K含量均表现出先降后升再下降的变化趋势,与另外3组的变化规律存在一定差异。
T2组在整个试验期间的全N含量平均值显著高于CK和T1组,分别高出8.3%、6.0%(P < 0.05)。虽然T3组全N含量平均值也分别高于CK和T1组3.7%、1.6%,但并未达到显著差异的程度(P>0.05)。T2与T3组全N含量平均值则无显著差异存在(P>0.05)。

|
图 2 不同水分条件下落羽杉幼苗实生土壤全氮、全磷和全钾含量的变化(±标准误) Figure 2 The change of total nitrogen, total phosphorus and total potassium in soils of T. distichum seedlings under different water treatment (±SE) |
各处理组土壤全P含量之间、全K含量之间分别均无显著差异存在(各处理组之间比较的P>0.05)。
2.3 土壤水解N、速效P和速效K的变化不同水分处理能显著影响到土壤水解N、速效P和速效K的含量(表 1)。随着水分处理的不断进行,CK组水解N含量表现出先升后降的变化趋势,在第3次测量时达到最大,这与另外3组的变化趋势有所不同。与CK组相比,T2组水解N含量始终处于CK组及其以上水平(第4次时T2与CK组比较,P>0.05),而T3组水解N含量则始终高于CK组0.1~0.3倍。T3组在整个试验期间的水解N含量平均值最高,分别显著高出CK、T1和T2组15.3%、5.4%和6.2% (P < 0.05)。与此同时,T1与T2组之间并无显著差异(T1与T2组:P>0.05),但均显著高于CK组水解N含量平均值(P < 0.05)。
T1组速效P含量始终处于CK组及其以上水平。T2、T3组在第1次测量时,速效P含量极显著地分别高于CK组58.4%、56.8%(P < 0.05),与在最后一次测量时反而极显著低于CK组47.2%、34.3%形成强烈对比(P < 0.05)。就整个试验期间的平均值而言,T2、T3组以及二者与CK组之间均无显著差异存在(各组间P>0.05)。相反,T1组的速效P含量平均值则极显著地分别高于另外3组(P < 0.05)。
T1组速效K含量也始终处于CK组及其以上水平(P>0.05),在整个试验期间的平均值最高,高于CK组达11.7%;这与T2、T3组以及二者与CK组之间均无显著差异形成对比(P>0.05)。

|
图 3 不同水分条件下落羽杉幼苗实生土壤水解氮、速效磷和速效钾的变化(±标准误) Figure 3 The change of alkali hydrolysable nitrogen, available phosphorus and available potassium in soils of T. distichum seedlings under different water treatment (±SE) |
相关性分析发现:土壤有机质含量与全N、全P以及水解N含量成极显著正相关,而pH值则与水解N、速效P含量成极显著负相关。全N与水解N、全P与速效P之间分别达到显著正相关,与全K和速效K之间并未表现出显著相关性形成对比。全P与全N、水解N之间分别表现出极显著正相关关系,而全P与全K之间却表现出极显著负相关性。速效K不仅与速效P表现出显著正相关性,还与全P之间表现出极显著正相关性。
|
|
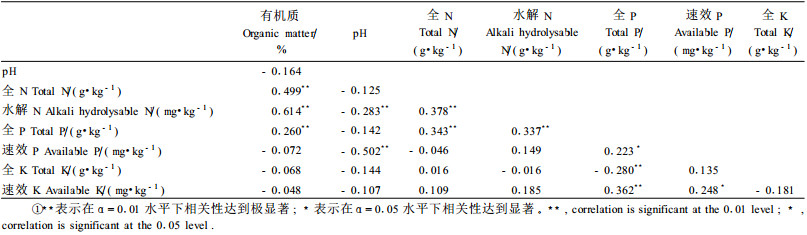
土壤养分是决定林木生长的重要因子(杨承栋等,2004;郝云庆等,2006)。通过与试验开始前的土壤营养元素含量初值比较发现,各营养元素含量均有不同程度的变化。各处理组在整个试验期间的土壤有机质含量平均值、pH平均值均显著增加,CK、T1、T2和T3组有机质含量比初值依次分别增加4.9%、11.0%、14.5%和15.3%,pH值比初始值依次分别提高15.0%、11.6%、15.4%和13.0%。这说明在落羽杉幼苗生长条件下,紫色土酸碱度将得到一定程度的调节,有机质含量会增加。谢锦升等(2002)研究发现,严重侵蚀红壤封禁管理后,随着地表植被的恢复,土壤有机质含量与pH值均增加,本研究结果与之一致。有机质含量的增加,与落羽杉幼苗枯枝落叶以及死亡根系进入到土壤中有关,还可能与土壤微生物种群和数量的增加有一定关系(Wilcke et al., 2002)。三峡水库消落区淹水后,酸性土壤的pH值升高,碱性土壤的pH值降低,并且二者都将接近于中性(pH 6.5~7.5)(石孝洪,2004),本试验结果也遵循这一规律。土壤pH值的改变,影响微生物种群、酶活性、有机质的分解,并且直接影响到养分的有效性(盛炜彤等,2003;安韶山等,2006)。落羽杉幼苗实生土壤pH值虽然增加,但基本上处于中性至微碱性条件,这恰好是大部分细菌和放线菌所适应的最佳酸碱度范围(金为民,2003),由此可以推测落羽杉幼苗实生紫色土中细菌和放线菌种群数量可能较大,养分转化速率可能较快。
各处理组土壤全N、全P含量平均值也均显著地高于对应的初值,这与各组全K含量并未与初始值表现出显著差异形成对比。但是各组水解N、速效P含量平均值则显著地低于各自对应的初始值,与速效K含量显著高于初始值截然相反。这很可能是因为落羽杉幼苗生长需要消耗较多的水解N、速效P,使得二者含量有所降低。李召青等(2005)通过对加勒比松幼龄林土壤肥力变化的研究发现,幼龄林土壤肥力有逐年下降的趋势,下降最快的是磷,其次是缓效钾,下降最慢的是速效氮。落羽杉幼苗土壤全N、全P含量平均值显著高于初值,与有机质含量的增加有较大关系,这也将有助于对土壤水解N、速效P的供给。由于土壤中的K与有机质没有关系(杨小波等,2000),其有效态主要为交换态,含量大小主要取决于土壤本身含量和土壤表面的聚集作用,故出现了与N、P元素不一致的变化结果。通过与土壤养分初值的比较发现,落羽杉幼苗恢复或改良土壤肥力的效果是比较明显的。这也进一步证实科学的植被恢复与植树造林能够有效地改良土壤、提高土壤肥力、保持水土(何斌等,2007;严代碧等,2007)。
在T3组条件下,土壤处于淹水缺氧环境中,氧化还原电位(Eh)降低,CO2分压增加,被固定、吸附的元素得以释放和移动(张甘霖等,1993)。与CK组相比,T3组有机质含量增加、pH值降低,说明水淹条件下落羽杉幼苗实生土壤受到酸性林溶程度有所增强。淹水条件下水解N含量增加,主要源于土壤有机质含量的增加,因为土壤N元素主要存在于有机质中,而且有机质含量与水解N含量存在着密切的相关性(杨承栋等,1986;Croker et al., 1955;Bormann et al., 1990)。蛋白酶和脲酶直接参与土壤中含N有机化合物的转化,两者活性强度可用来表征土壤N元素供应程度(万福绪等,2006)。因此,T3组条件下水解N含量的增加还可能与蛋白酶和脲酶的活性增强有一定关系。土壤全N、全P、全K含量在淹水条件下与CK组相比均未出现显著差异,表明淹水条件下土壤供应植物必须的三大营养元素容量并未发生显著变化;速效P、速效K与CK组相比也未表现出显著差异性,证实土壤供应P、K元素的强度也并未因水淹条件而发生显著变化。
在T2组条件下,土壤有机质、全N、水解N含量均分别显著高于CK组,而pH值、全P、全K、速效P、速效K含量与CK组相比均未出现显著差异。这与T3组土壤养分含量的变化有所不同。T2组维持与CK组同样的pH值水平,显示出饱和水分条件并没有对土壤造成较大淋溶作用,对土壤酸性环境的形成并不显著(刘友良,1992)。T2组全N与水解N含量同时增加,可能与pH值维持在CK组正常水平关系密切。T2组全N含量增加,使得饱和水土壤的供N容量增大,为水解N含量的增加提供可能。
与T2、T3组均有所差异,T1组的有机质、水解N、速效P与速效K含量均显著高于CK组,pH值显著低于CK组,而全N、全P、全K含量与CK组相比却未出现显著差异。在轻度干旱条件下,土壤氧化还原能力较强,土壤转化酶、脲酶、蛋白酶以及酸性磷酸酶活性均较强。转化酶活性过低,要影响到土壤中各种有机物的分解与转化,从而直接或间接地要影响到各种代谢活动的正常进行;土壤转化酶活性的高低,通常是森林土壤生产力的重要标志(周礼恺,1987;安韶山等,2006)。土壤酸性磷酸酶的酶促作用能加速土壤有机磷的脱磷速度,提高磷素的有效性。在T1组条件下,土壤供应养分的容量保持在正常水平,而养分供应的强度提高,说明轻度干旱水分条件有助于落羽杉幼苗实生紫色土的营养元素释放。
石孝洪等(2004)通过对三峡库区消落带土壤磷吸附特征的模拟研究,发现土壤淹水后,冲积土和紫色土释放磷的风险大于黄壤,磷的有效性提高,促进了磷的释放。这是在没有实生植物生长条件下,不同土壤类型之间比较所得到的结论。而本研究通过模拟三峡库区消落带土壤淹水变化条件,盆栽落羽杉幼苗,对植物实生土壤进行测定,发现T3组土壤磷的有效性在整个试验期间的平均值与CK组相比并未出现显著差异。不同的试验条件得到了有关磷的有效性的不同结论。本试验结果显示:水淹条件下的全P含量平均值与轻度干旱条件下相比差异不显著,但轻度干旱条件下的速效P含量平均值则显著高于水淹条件,与石孝洪等(2004)测定消落区旱地与水田土壤所得试验结果一致。张金洋等(2004)在没有植物生长条件下,模拟研究消落区在水淹条件下的土壤性质变化状况,发现土壤中Zn、Cu、Pb、Cd、Cr、Hg、As的含量都有不同程度的降低,其中以Hg降低较为显著,而有效态的Zn、Cu、Pb、Cd、Te和Mn的含量大部分呈增加趋势。可以推测,如果有植物生长的实生土壤,上述微量元素含量在水淹状态下的变化也可能与之有所差异。
落羽杉对土壤具有一定的改良作用,通过对土壤的改良可以进一步促进自身的生长发育。目前,三峡库区内土壤退化严重,土壤肥力在逐步下降;随着侵蚀程度的增加,土壤中N、P、K及有机质将普遍缺乏(杜榕桓等,1994)。因此,在三峡库区消落带适度营造落羽杉对于恢复库区植被、改良土壤状况具有积极作用。
安韶山, 黄懿梅. 2006. 黄土丘陵区柠条林改良土壤作用的研究. 林业科学, 42(1): 70-74. |
杜榕桓, 史德明, 袁建模, 等. 1994. 长江三峡库区水土流失对生态与环境的影响. 北京: 科学出版社, 81-86.
|
杜荣骞. 2003. 生物统计学. 2版. 北京: 高等教育出版社, 104-116.
|
郝云庆, 何丙辉, 李旭光. 2006. 巫溪县红池坝不同植被恢复阶段土壤养分评价. 西南农业大学学报:自然科学版, 28(1): 149-153. |
何斌, 贾黎明, 金大刚, 等. 2007. 广西南宁马占相思人工林土壤肥力变化的研究. 林业科学, 43(5): 10-16. |
胡新生, 王世绩. 1998. 树木水分胁迫生理与耐旱性研究进展及展望. 林业科学, 34(2): 77-88. DOI:10.3321/j.issn:1001-7488.1998.02.011 |
胡哲森, 许长钦, 傅瑞树. 2000. 锥栗幼苗对水分胁迫的生理响应及6-BA的作用. 福建林学院学报, 20(3): 1-4. |
金为民. 2003. 土壤肥料. 北京: 中国农业出版社, 47-51, 148-152.
|
李昌晓, 钟章成. 2005. 模拟三峡库区消落带土壤水分变化条件下落羽杉与池杉幼苗的光合特性比较. 林业科学, 41(6): 28-34. DOI:10.3321/j.issn:1001-7488.2005.06.005 |
李其林, 黄昀, 刘光德, 等. 2004. 三峡库区主要土壤类型重金属含量及特征. 土壤学报, 41(2): 301-304. DOI:10.3321/j.issn:0564-3929.2004.02.022 |
李召青, 陈淑贤, 周毅, 等. 2005. 加勒比松造林后土壤肥力变化研究. 广东林业科技, 21(4): 30-33. DOI:10.3969/j.issn.1006-4427.2005.04.008 |
刘友良. 1992. 植物水分逆境生理. 北京: 农业出版社, 154-160.
|
盛炜彤, 杨承栋, 范少辉. 2003. 杉木人工林的土壤性质变化. 林业科学研究, 16(4): 377-385. DOI:10.3321/j.issn:1001-1498.2003.04.001 |
石孝洪. 2004a. 三峡水库消落区土壤磷素释放与富营养化. 土壤肥料, (1): 40-44. |
石孝洪, 魏世强, 谢德体, 等. 2004b. 三峡水库消落区土壤磷吸附特征. 西南农业大学学报:自然科学版, 26(3): 331-335. |
万福绪, 杨东. 2006. 苏北海堤杉木杨树混交林林木生长及土壤肥力研究. 南京林业大学学报:自然科学版, 30(2): 43-46. |
汪贵斌, 曹福亮. 2002. 落羽杉抗性研究综述. 南京林业大学学报:自然科学版, 26(6): 78-82. |
谢锦升, 杨玉盛, 陈光水, 等. 2002. 严重侵蚀红壤封禁管理后土壤性质的变化. 福建林学院学报, 22(3): 236-239. DOI:10.3969/j.issn.1001-389X.2002.03.011 |
严代碧, 岳永杰, 宿以明, 等. 2007. 岷江上游干旱河谷辐射松人工幼林营养元素的研究. 南京林业大学学报:自然科学版, 31(1): 31-36. |
杨承栋, 张万儒. 1986. 卧龙自然保护区森林土壤有机质的研究. 土壤学报, 23(1): 30-39. |
杨承栋, 张小泉, 魏以荣, 等. 2004. 浙江省丽水不同前茬杉木林土壤性质与林木生长. 土壤学报, 41(6): 954-961. DOI:10.3321/j.issn:0564-3929.2004.06.018 |
杨刚. 2004. 三峡库区受淹土壤污染物释放量的试验研究. 水土保持学报, 18(1): 111-114. DOI:10.3321/j.issn:1009-2242.2004.01.029 |
杨小波, 胡荣贵. 2000. 热带滨海沙滩上森林植被的组成成分与土壤性质的研究. 生态学杂志, 19(4): 6-11. DOI:10.3321/j.issn:1000-4890.2000.04.002 |
岳庆玲, 常庆瑞, 刘京, 等. 2007. 黄土丘陵沟壑区不同人工林地土壤肥力变化研究. 干旱地区农业研究, 25(3): 100-104. DOI:10.3321/j.issn:1000-7601.2007.03.021 |
张甘霖, 龚子同. 1993. 淹水条件下土壤中元素迁移的地球化学特征. 土壤学报, 30(4): 355-365. DOI:10.3321/j.issn:0564-3929.1993.04.013 |
张金洋, 王定勇, 石孝洪. 2004. 三峡水库消落区淹水后土壤性质变化的模拟研究. 水土保持学报, 18(6): 120-123. DOI:10.3321/j.issn:1009-2242.2004.06.029 |
中国土壤学会农业化学专业委员会. 1983. 土壤农业化学常规分析方法. 北京: 科学出版社, 100-115.
|
周礼恺. 1987. 土壤酶学. 北京: 科学出版社, 237-239.
|
Bormann B T, Sidle R C. 1990. Change in productivity and distribution of nutrients in a chrono-sequence at Glacier bay, national park, Alaska. Journal of Ecology, 18: 561-578. |
Bragina T V, Martinovich L I, Rodionova N A, et al. 2001. Ethylene-induced activation of xylanase in adventitious roots of maize as a response to the stress effect of root submersion. Applied Biochemistry and Microbiology, 37(6): 618-621. DOI:10.1023/A:1012363319758 |
Croker R L, Major J. 1955. Soil development in relation to vegetation and surface age at Glacier bay, Alaska. Journal of Ecology, 43: 427-448. DOI:10.2307/2257005 |
Ponnamperuma F N. 1972. The chemistry of submerged soil. Advances in Agronomy, 24: 29-36. DOI:10.1016/S0065-2113(08)60633-1 |
Randall C K, Duryea M L, Vince S W. 2005. Factors influencing stump sprouting by pondcypress (Taxodium distichum var. nutans(Ait.)Sweet). New Forests, 29: 245-260. DOI:10.1007/s11056-005-5652-4 |
Wilcke W, Lilienfein J. 2002. Biogeochemical consequences of the transformation of native Cerrado into Pinus caribaea plantations in Brazil—Biogeochemical cycling in Cerrado and Pinus caribaea forests. Plant and Soil, 238: 175-189. DOI:10.1023/A:1014421305608 |