 2008, Vol. 44
2008, Vol. 44文章信息
- 张文辉, 卢彦昌, 周建云, 张晓辉, 史小华.
- Zhang Wenhui, Lu Yanchang, Zhou Jianyun, Zhang Xiaohui, Shi Xiaohua.
- 巴山北坡不同干扰条件下栓皮栎种群结构与动态
- Population Structure and Dynamics of Quercus variabilis in Different Habitats on Northern Slope of Bashan Mountain
- 林业科学, 2008, 44(7): 11-16.
- Scientia Silvae Sinicae, 2008, 44(7): 11-16.
-
文章历史
- 收稿日期:2006-12-25
-
作者相关文章


植物种群结构和动态是植物个体生存能力与外界环境相互作用的结果。在一定区域内,植物群落向与当地气候和土壤相适应的稳定方向演替。在持续外界干扰下,植物群落可能偏离演替方向形成偏途顶极。在相似生境条件下,对照研究封禁和持续干扰条件下地带性森林群落建群种种群结构与动态,对阐明群落更新演替及维持生态系统具有重要意义(Manuel,2000)。
栓皮栎(Quercus variabilis)为落叶乔木,广泛分布于我国22个省区(张文辉等,2002a),是中国暖温带落叶阔叶林林区地带性植被的主要建群种,也是软木、栲胶、食用菌、天麻生产资源树种(傅焕光等,1986)。由于用途广泛,屡遭破坏,目前大部分栓皮栎林已经沦为残败次生林。陕西秦巴山不少地区将栓皮栎林作为薪炭林经营,多年连续砍伐导致种群灌木化(韩照祥等,2005)。外界干扰导致栓皮栎种群产生哪些变化?针对大面积惨败次生林,通过什么途径才能恢复残败次生林、维持栓皮栎林分更新能力?这些是当前生产实际中的突出问题。
目前对巴山地区栓皮栎林的研究主要集中在群落物种多样性、地理分布、分布格局等方面(李林等,2006;张文辉等,2002b;韩照祥等,2005),有关外界干扰与种群结构和幼苗更新方面的研究很少。巴山北坡岚皋县大部分地区的栓皮栎林在20世纪50年代末曾被皆伐。随后,远离村落地区的林分属于集体林或国有林,基本处于封禁状态,而距离村落较近的林地则被划为薪炭林,由于砍耳棒(培养食用菌木耳或香菇的木棒)和薪柴,处于持续干扰状态下。经过50多年的发育,2种生境的林分外貌、建群种种群结构、更新状态出现了明显差异。本研究通过在2种林地的相似生境设置样地,调查栓皮栎的种群结构、数量动态、幼苗更新和空间分布格局,目的是阐明外界干扰对栓皮栎种群动态和更新状况的影响,探索种群维持与恢复的途径,为次生林的恢复及管理提供依据。
1 研究区概况研究区位于陕西省安康市岚皋县(31°42′—33°49′ N,108°01′—110°01′ E),属于巴山北坡,北亚热带气候,年降水量1 017 mm,年均气温15 ℃。土壤为山地褐土或山地棕壤土,地带性植被为亚热带常绿阔叶林与落叶阔叶林相间分布(雷明德,1999)。栓皮栎林是当地主要森林类型,主要分布在海拔500~1 700 m处,乔木层郁闭度为0.5~0.9,伴生树种主要有麻栎(Quercus acutissima)及马尾松(Pinus massoniana)等,高度为8~12 m;林下灌木盖度为0.3~0.5,高度为0.5~1.5 m,主要有胡枝子(Lespedeza bicolor)、黄栌(Continus coggygria)、榛子(Corylus heterophylla)及陕西荚NFDA2 (Viburnum schensianum)等;草本层盖度为0.4~0.6,高为10~40 cm,以禾本科的白茅草(Imperata cylindrical var. major)、白羊草(Bothrichloa ischaemum)和莎草科的苔草属(Carex)植物为主。
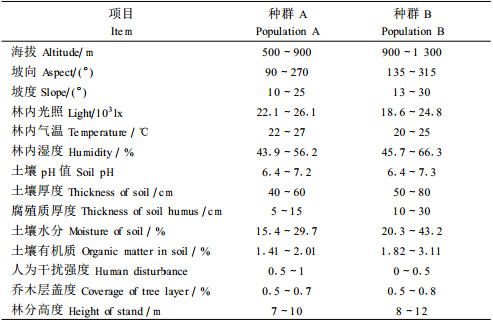
2 研究方法 2.1 样地设置与调查内容研究所选择的栓皮栎林地在1958—1959年曾进行过皆伐。此后,一部分林地离居民点比较近(城关镇周边地区),海拔500~900 m,四周往往与荒草地、农田交错分布,人为活动频繁,受干扰强度大,20世纪80年代以后由农户承包,作为耳林(培养食用菌木耳的专用林)或薪炭林经营,作为种群A。另一部分林地海拔900~1 300 m,远离居民点,属于集体林或国有林,由林场或保护区(现为南宫山森林公园)管理,地理位置特殊,近50年来基本处于封禁状态,作为种群B。2006年7—8月,在2个种群林分发育较好的地段各设置6块20 m×20 m的样地,其中在南向、东南向与西南向山坡中部各设样地2块。通过调查、统计,得到2个种群的概况表(表 1)。测定每一个体(包括幼苗、幼树)的坐标定位(以样地一个角为原点,与坡面等高线平行边为x轴,垂直边为y轴)、高度、胸径(幼苗幼树的基径)、冠幅及枝下高等(张文辉等,2005)。
|
|
参照高贤明等(2001)对辽东栎(Quercus wutaishanica)的研究结果,将高度<2 m和胸径(DBH)<2 cm的栓皮栎视为幼苗(树)。调查中将独立生长基部无伐桩痕迹的个体均确定为实生苗,将基部有明显伐桩痕迹的幼苗和大树基部的萌苗,确定为萌生苗。
2.3 径级结构与静态生命表编制以个体的高度(苗期)及径级代替龄级来反映栓皮栎的种群结构(王巍等,1999)。将幼龄个体划分为幼苗(高度≤40 cm)、小幼树(高度为40~100 cm)和大幼树(高度>100 cm,DBH<2 cm)3个级别,径级分别为Ⅰ,Ⅱ和Ⅲ,然后从DBH 2 cm起,每增加2 cm为一个径级,依次编号为Ⅳ,Ⅴ,Ⅵ…,分别统计各径级株数并绘制种群径级结构图、编制静态生命表,根据生命表中的标准化存活数和径级绘制存活曲线(江洪,1992)。
2.4 分布格局分析以样地调查中栓皮栎种群个体坐标值为基础数据,以5 m×5 m作为测算单元,采用相邻格子法进行格局分析。采用Morisita分布指数(Iδ)、方差均值比(V/X)和聚块性指数(m*/m)作为种群分布格局判定依据(张文辉等,2003;赖江山等,2006;张金屯,2004)。
2.5 主成分分析利用SPSS统计软件,以各样地相对独立的环境因子作为基础(表 1),建立原始矩阵,进行主成分分折,确定环境因子中影响栓皮栎种群发育的主要因子。
3 结果与分析 3.1 种群径级结构及动态从图 1可以看出,栓皮栎种群在2种生境条件下都具有一定数量的幼龄个体,但种群A幼苗和幼树数量不足,种群密度小。种群B的径级结构基本上属于倒J字形分布,幼苗、幼树个体数量丰富,种群密度大。种群B更新状况明显强于种群A。两者差异主要来源于水、热、土壤和外界干扰因素的影响。种群A分布于海拔700~900 m处,村民砍柴、砍耳棒、剥栓皮等外界干扰严重,林下地面枯枝落叶少,腐殖质层薄,土壤贫瘠,种群幼龄个体数量少,并以伐桩萌芽幼苗占优势,种群总体呈现衰退势态。种群B分布于海拔900~1 300 m处,远离村庄,人为干扰较少,生境条件有利于栓皮栎种子的萌发、定居和萌生苗的生长,幼龄个体数量丰富,并以实生苗为主,种群呈现持续发育势态。

|
图 1 A、B种群径级结构 Figure 1 Diameter structure of population A and B |
种群幼龄个体数量是种群更新的基础,不同起源对种群遗传多样性维持意义重大。为了阐明不同生境栓皮栎种群更新主体,本文统计了幼苗、小幼树和大幼树个体。从图 2可看出,对于种群A,实生苗在幼苗、小幼树和大幼树中的比例分别为58.57%、31.01和17.70%;对于种群B,实生苗在幼苗、小幼树和大幼树中的比例分别为88.56%、70.12%和56.19%。很明显,种群A萌生苗占优势,而种群B实生苗占优势。

|
图 2 幼龄个体组成 Figure 2 Young individual composition |
种群A和B的共同之处表现为从幼苗到小幼树、大幼树的发育过程中,实生种群的比例在减小,而萌生种群的比例在增大。栓皮栎实生苗在早期主要是地下根系的生长,而地上枝叶生长非常缓慢。萌生苗根系庞大,在初期地上部分生长迅速。与相同年龄的实生苗相比,萌生苗在空间分布和对资源利用等方面均占有明显的优势,而实生幼苗处于不利地位,由于竞争导致死亡,其比例也不断减小(Guama et al., 2006)。实生苗是由种子发育而成,对于维持种群遗传多样性极为重要。萌生苗主要通过伐桩萌生产生,其遗传基础与亲本完全一致,随着砍伐次数增加和生理衰老,种群也会出现相应的衰退症状。要使种群长期稳定持续发育,应尽量促进实生苗发育(McEvoy et al.,2006)。
3.3 种群生命表和存活曲线静态生命表能够反映种群在与环境因素相互作用过程中的个体生死过程,对阐明种群生活史的脆弱环节很重要。从表 2可以看出,在A种群中,Ⅰ径级死亡率最高,在B种群中,Ⅱ径级死亡率最高。这2个发育阶段,可能就是种群生活史中的脆弱阶段,应该加强保护。在Ⅲ与Ⅳ龄级,2个种群期望寿命最高,说明这2个龄级的栓皮栎种群生存能力最强。2个种群死亡量(率)在某些龄级出现负值,可能与人为干扰有关。
|
|

栓皮栎2个种群存活曲线均属于DeeveyⅢ型(图 3),幼龄期死亡率高,是种群生活史的脆弱环节。这与张文辉等(2003)有关秦岭北坡栓皮栎种群存活曲线基本一致。保护较好的种群B基本代表了这个特点,在度过大幼树阶段以后,个体存活数量基本稳定。当一般林木个体进入主林层后,个体死亡率下降到较低水平。由于种群A受外界干扰严重,出现Ⅲ与Ⅳ径阶个体存活数量明显低于Ⅴ,Ⅵ与Ⅶ径级的现象,说明外界干扰已经影响到种群萌芽力和实生苗的正常发育,保护好幼龄阶段的苗木是种群恢复的关键。

|
图 3 种群存活曲线 Figure 3 Survival curve of populations |
在发育不同阶段分布格局反映了种群占据空间的能力与生存策略调整。表 3说明2个种群在幼苗、小幼树、大幼树向大树发育过程中都呈聚集分布,但聚集强度随年龄增加逐渐减弱。种群A在幼龄阶段聚集强度弱,但在发育过程中聚集强度下降缓慢,而种群B与之相反。两者差异主要来源于人为干扰。在种群B中,个体多为实生起源、种子堆积、林下生境异质等导致聚集分布。在种群A中,伐桩萌苗占主导地位,伐桩分布格局决定了幼龄个体分布,所以种群聚集强度低是正常现象。
|
|

选取表 1中相对独立的环境因子进行主成分分析(表 4)。由于前2个主成分的累积贡献率已经达到83.49%,故本文仅对前2个主成分进行综合分析。第一主成分中,人为干扰的负荷量为-0.959,是影响栓皮栎种群结构最主要的负面影响因子;海拔高度的负荷量是0.908,随海拔升高人为干扰强度减小,降雨量增加,对种群发育产生正面影响。此外,土壤水分含量(0.948)、腐殖质厚度(0.937)、土壤有机质含量(0.909)和土壤厚度(0.877)的负荷量也较高,这些土壤因素对种群发育形成具有重要的正面影响。在第二主成分中,林内光照强度(0.876)、林内温度(0.840)是影响种群发育的主要因素。林内光照强度及温度高,有利于栓皮栎生长。因此,减少人为干扰,增加林内光照、温度、土壤水分、腐殖质层厚度和土壤有机质有利于栓皮栎种群良性发展。
|
|

保护较好的B种群径级结构、生命表、分布格局等指标都显示出种群呈持续发育状态。而A种群长期受到持续干扰,种群密度低,幼苗数量不足,熟生苗比例较低,种群具有不稳定特征。McEvoy(2006)研究认为持续放牧干扰导致Quercus robur种群密度降低,实生苗缺乏,而不放牧或者很少放牧的天然林地有性实生苗占绝对优势,群落相对稳定。本研究对影响种群结构的关键因子进行主成分分析,结果也表明,人为干扰是最重要的不利因素。因此,采取封禁措施,减少林地干扰,可发挥栓皮栎自然恢复潜力,在有条件时也可进行人工抚育以促进恢复。
在次生林恢复中,萌生幼苗发挥了重要功能。但植物萌芽力是有限度的,连续砍伐会导致伐桩萌芽力降低,A种群Ⅲ与Ⅳ径阶个体数量明显降低可能与此有关。Roy等(2006)认为栎类无性繁殖短期恢复群落生态功能的作用巨大,但从森林持续稳定角度来看,更要注意保护实生幼苗,以维持种群丰富的遗传多样性。
外界环境因素对种群径级结构、幼龄个体组成和分布格局有重要影响。生命表和存活曲线分析结果表明,栓皮栎幼龄阶段是种群生活史中的脆弱环节。因此,未来经营管理中,应围绕抚育栓皮栎幼苗这一目标进行环境调控,保证足够数量的幼苗,保障种群持续发展。在高海拔地区,应适时适度地间伐非目的树种,促进栓皮栎个体发育。在低海拔地区,应轮封轮放,既要抚育萌生苗,又要培育结果母树,促进实生幼苗发育。
傅焕光, 于光明. 1986. 栓皮栎栽培与利用. 北京: 中国林业出版社, 1-20.
|
高贤明, 王巍, 杜晓军, 等. 2001. 北京山区辽东栎林的径级结构、种群起源及生态学意义. 植物生态学报, 25(6): 673-678. DOI:10.3321/j.issn:1005-264X.2001.06.005 |
韩照祥, 朱惠娟, 张文辉, 等. 2005. 不同地区不同尺度下栓皮栎种群的空间分布格局. 西北植物学报, 25(6): 1216-1221. DOI:10.3321/j.issn:1000-4025.2005.06.029 |
江洪. 1992. 云杉种群生态学研究. 北京: 中国林业出版社, 8-50.
|
赖江山, 张谧, 谢宗强. 2006. 三峡库区常绿阔叶林优势种群的结构和格局动态. 生态学报, 26(4): 1073-1079. DOI:10.3321/j.issn:1000-0933.2006.04.013 |
雷明德. 1999. 陕西植被. 北京: 科学出版社, 163-164.
|
李林, 黄忠良, 张海忠, 等. 2006. 陕西省栓皮栎群落物种多样性的空间异质性. 福建林学院学报, 26(1): 63-68. DOI:10.3969/j.issn.1001-389X.2006.01.015 |
王巍, 刘灿然, 马克平, 等. 1999. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 41(4): 425-432. |
张金屯. 2004. 数量生态学. 北京: 科学出版社, 243-266.
|
张文辉, 卢志军. 2002a. 栓皮栎种群生物学生态学和地理分布研究. 西北植物学报, 22(5): 1093-1101. |
张文辉, 卢志军, 李景侠, 等. 2002b. 陕西不同林区栓皮栎种群空间分布格局及动态的比较研究. 西北植物学报, 22(3): 476-483. |
张文辉, 卢志军, 李景侠, 等. 2003. 秦岭北坡栓皮栎种群动态的研究. 应用生态学报, 14(9): 1427-1432. DOI:10.3321/j.issn:1001-9332.2003.09.006 |
张文辉, 许晓波, 周建云, 等. 2005. 濒危植物秦岭冷杉种群数量动态. 应用生态学报, 16(10): 1799-1804. DOI:10.3321/j.issn:1001-9332.2005.10.001 |
Guama J, Linera G. 2006. Edge effect on acorn removal and oak seedling survival in Mexican lower montane forest fragments. New Forests, 31: 487-495. DOI:10.1007/s11056-005-2015-0 |
Manuel C M. 2000. Ecology: concepts and application. Beijing: Science Press.
|
Crawley M J, Long C R. 1995. Alternate bearing, predator satiation and seedling recruitment in Quercus robur. Journal of Ecology, 83: 683-696. DOI:10.2307/2261636 |
McEvoy P, McAdaml J, Losada M, et al. 2006. Tree regeneration and sapling damage of pedunculate oak Quercus robur in a grazed forest in Galicia, NW Spain: a comparison of continuous and rotational grazing systems. Agroforestry System, 66: 85-92. DOI:10.1007/s10457-005-2916-0 |
Roy B, Lockhart J, Chambers L. 2006. Cherrybark oak stump sprout survival and development five years following plantation thinning in the lower Mississippi alluvial valley, USA. New Forests, 31: 22-27. |