 2008, Vol. 44
2008, Vol. 44文章信息
- 周亮, 刘盛全, 朱永侠, 黄振英, 邵卓平.
- Zhou Liang, Liu Shengquan, Zhu Yongxia, Huang Zhenying, Shao Zhuoping.
- 马尾松生长性状与生长应力的关系
- Relationship between Growth Traits and Growth Stress of Masson Pine
- 林业科学, 2008, 44(6): 101-108.
- Scientia Silvae Sinicae, 2008, 44(6): 101-108.
-
文章历史
- 收稿日期:2007-07-04
-
作者相关文章



高生长应力和应力分布不均是目前人工林速生材普遍存在的木材缺陷之一(Archer,1987)。由于生长应力的存在和分布的不均,生长应力大的立木刚伐下时就容易产生轮裂、脆心、脆性和压伤等缺陷。在原木锯解等过程中木材内部应力的平衡被打破,应力释放后产生弓弯、边弯、扭曲和夹锯等,给制材行业带来很大的经济损失(Yang et al., 2002;Chafe,1979),所以各国木材科学研究学者一直在探讨和研究树木生长应力的影响因子、形成机制和调控方法(Wang,1977;Okuyama et al., 1987;)。关于生长应力(growth stress)的形成原因和机制目前还没有统一定论,主要有3种不同的假说,分别是木素膨胀假说(Watanabe,1965;Boyd,1972)、纤维拉力假说(Bamber,1979;1987)、统一假说(Yamamoto,1998),但是国内外学者一致认为生长应力形成于树木次生长过程当中。由于木材内部生长应力是在木材次生生长过程中形成的,生长应力与立木次生生长形成的生长性状以及生长的立地条件有着一定联系,所以国外学者一直在尝试将这两者联系起来研究,为通过人工林栽培措施和林木选育的方法来降低立木生长应力提供一定的理论依据(Wang,1972;Okuyama et al., 1981;Ferrand,1982;Saurat et al., 1976;Polge,1981;Malan et al., 1987),并可以在立木状态下对生长应力的大小进行初步预测评估。如Saurat等(1976)对法国不同省份栽植的欧洲水青冈(Fagus sylvatica)轴向表面生长应力和应变研究中发现,复层林的平均应变、应力值和应变变化范围都比单层林少40%;Polge(1981)对100年以上欧洲水青冈生长应力钻取生长锥,通过观察其直径变化研究生长应力得出高强度的间伐能降低立木生长应力,并且认为树木冠幅越大,生长应力值越小;Malan等(1987)采用计算伐倒木断头开裂来估算在南非栽植的巨桉(Eucalyptus grandis)生长应力,并以此为依据研究生长应力大小与生长速度和冠幅之间的关系,但研究结果与以上学者研究结论相驳,多元回归得出的结论认为生长速度和冠幅只能对生长应力变异中的很少一部分进行解释,并据此推断栽培措施不能有效降低生长应力;我国学者胡继青等(2000)对雷州1号桉(Eucalyptus leizhou No.1)、柠檬桉(E. citriodora)和尾叶桉(E. urophylla)立木的轴向生长应变时得出,树高对生长应变的影响可以忽略,雷州1号桉与柠檬桉立木生长应变与胸径相关,前者呈正相关,后者呈负相关,尾叶桉为不相关;费本华等(2004)研究人工林桉树立木生长应变与立木胸径关系时也得出,由于桉树种源不同、树龄不同,生长应变与胸径之间关系也存在较大差异。可见由于测试方法、树种和采集地不同,得出的结论也不一致。
马尾松(Pinus massoniana)是我国丰富的木材资源之一,遍布南方15个省(区),是重要的用材林树种和造林先锋树种(安徽农学院林学系,1980)。在速生人工林马尾松树木中存在着一定的生长应力,并伴有应压木的产生,对马尾松木材的加工利用造成负面影响(林金星等,1993;黄振英,2004),然而对人工速生马尾松立木生长应力与生长性状的研究还未见报道。本文尝试以2种不同林型下3个不同树干形态的人工速生马尾松为研究对象,选择3块不同立地条件的样地,通过对其轴向表面生长应力和生长性状的测定,分析两者之间的关系,寻找影响马尾松表面轴向生长应力的主要因子,以期为通过栽培措施和林木育种降低马尾松人工速生林生长应力的方法提供理论依据。
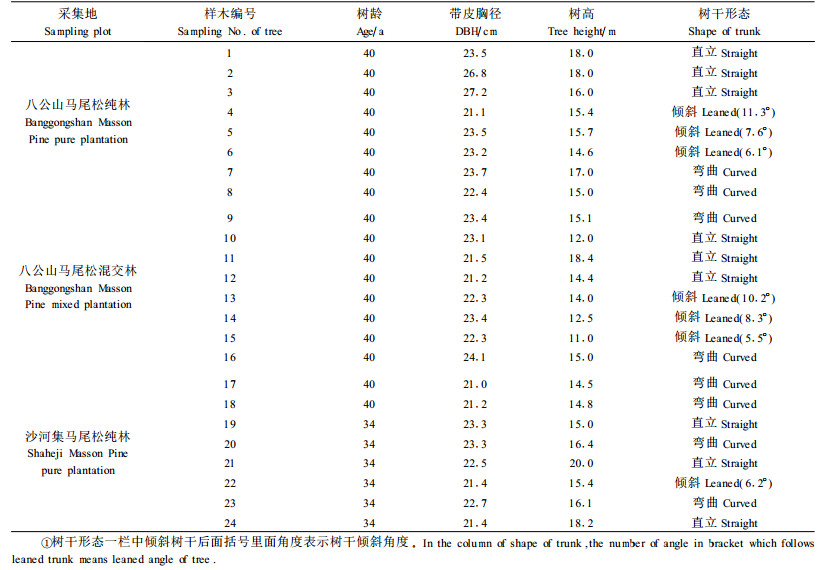
1 材料与方法 1.1 采集地与样木选取3块不同立地条件的马尾松试验林:第1块是位于安徽省寿县八公山林场马尾松人工林纯林;第2块是同一林场马尾松与麻栎(Quercus acutissima)混交林;第3块是安徽省滁州市沙河集总场大柳分场马尾松人工林纯林。八公山林场马尾松纯林位于丘陵地的阴坡,坡度为30°,混交林为阳坡,坡度为50°,土壤类型均为黄棕壤,年均气温15~20 ℃,年均降水量916 mm; 沙河集马尾松人工林纯林位于丘陵地的阳坡,坡度为5°,土壤类型为黄棕壤,年均气温为14.4~16.6 ℃,年均降水量1 031.2 mm。在八公山林场内分别按照各自小班数据,选取马尾松纯林和混交林各9株(通直、倾斜、弯曲3种不同树形各3株),共18株。其中,对于弯曲树干的样木要求在胸高处附近出现弯曲。沙河集林场内选取样木6株(通直的3株,倾斜的1株,弯曲的2株),3块试验地共完成24株马尾松立木生长应力测试。所选样木生长性状情况如表 1所示,弯曲和倾斜树干形态草图如表 2所示。
|
|
|
|

常规方法测定树高、胸径、树龄;以铅垂线为对照,描绘第一活枝以下的树干形态草图,并以此为依据区分树干形态;树干倾斜角度为1.3 m以下树干与铅垂线的夹角,通过测定三角形三边长度,利用勾股定律计算得知。
1.2.2 测试点的选择为了有效分析周向不同测试点与生长应力之间关系,本文共采用2种不同测试点选择方法,一种为八等分法(沙河集林场马尾松生长应力研究),另一种为四等分法(八公山林场马尾松生长应力研究)。测试点的选择如图 1所示,测试点编号顺序从倾斜树干下部、弯曲树干凸处、直立树干北方向到从倾斜树干上部、弯曲树干凹处、直立树干南方向沿着直径方向逐渐增加。

|
图 1 各测试点在树干外围位置及其编号示意图 Figure 1 Sketch map for illustrating the relative position and serial No. of testing points around trunk periphery 括号内数字为四等分法所采用编号 The number in the bracket is serial No. of testing points of the quartering method. |
树木生长应力往往很难直接被测得,目前国内外主要通过对立木生长应变的测定来计算生长应力,计算公式为σ≈-E·ε(σ为生长应力,ε为生长应变,E为弹性模量)。本文选用法国热带木材研究中心研制的CIRAD-Forêt生长应变仪对立木轴向表面生长应变进行测试(Fournier et al., 1994),其原理是通过在立木树干上钻孔使生长应力局部释放,随后通过测定钻孔前后2个金属钉之间沿纤维方向上位移变化值δ来估计立木轴向生长应变,计算公式为ε=-Φ·δ(ε为轴向表面生长应变,Φ为一常数,取决于孔的直径、测量距离、测试材料的弹性模量以及泊松比,对于马尾松具体取值未知),然后再由生长应变计算生长应力,其公式为σ≈E·Φ·δ。对于一个特定材料,如马尾松立木,公式中E·Φ为常数项,有具体数值且为正值,所以用此方法测定立木表面生长应力,只需要测定δ即可,并且根据公式明显看出,δ与σ为线性正相关关系,因此在通常情况下为了研究的简便,在对立木轴向表面生长应力进行测定时,不进行E和Φ的测定,不计算出具体σ,直接用尺寸变化量δ作为轴向表面生长应力的指示参数来表征轴向表面生长应力,称为生长应力指示值(growth stress indicator,简写成GSI)(Fournier et al., 1994;Gerard et al., 1995;Clair et al., 2003)。
野外测试时,首先用铲子和斧子除去每个等分点上的树皮(20 cm高×10 cm宽),在剥去树皮的同时一定不能损伤到裸露的新鲜木质部,然后再依次在每个等分点上固定纵向应变测试装置,通过锥钻法释放纵向生长应力,手钻锥入深度大约在15~20 mm左右,直到读数基本稳定后记下数值,即生长应力指示值GSI。
1.2.4 数据处理方法选用SPSS统计软件对所得生长应力指示值进行多元方差分析、相关分析,计算偏相关系数。
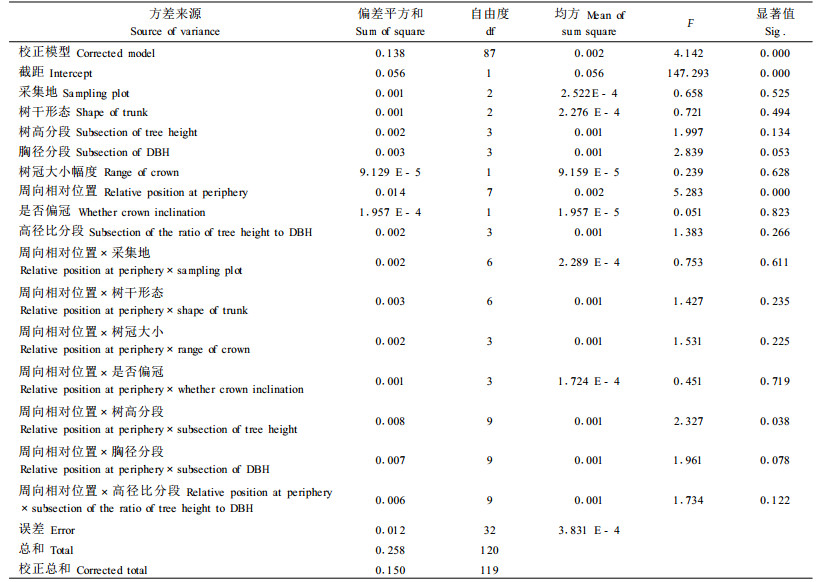
2 结果与分析 2.1 生长性状对马尾松树干表面轴向GSI的影响影响因变量表面轴向GSI的自变量可以分为2类:一类为定性描述的变量,如周向相对位置(在周向上测试点相对位置,用从数字1~8表示)、树干形态、冠型大小、是否偏冠,本文中统称为定性自变量;另一类为定量描述的生长性状,如树高、胸径、高径比、倾斜角。对于此类连续的自变量,在方差分析之前调用SPSS软件中的可视化分段功能,将每组数据中插入3个等分点,即把原来每组数据转变成4个序数值进行方差分析,所得自变量本文中统称为定量自变量。为了降低周向相对位置对其他变量方差分析结果的影响,将所有变量主效应和周向相对位置和其余变量之间的二维交互效应列入多因素方差分析模型,所得结果如表 3所示。为进一步分析各自变量对GSI的影响,在方差分析的同时,采用S-N-K法对周向相对位置、树干形态、采集地分别进行两两比较,结果如表 4所示。
|
|
|
|

由表 3可以看出,多因素方差分析模型在0.001水平上显著。R2是以GSI为因变量进行多元线形回归方程的决定系数,它的数值表示分析模型已经解释了因变量总变异的多少,即多因素方差分析中所指定模型已经解释总变异的93.4%。由方差模型分析得出:主效应周向相对位置对GSI影响在0.001水平上显著;周向相对位置和树高分段的二元交互效应对GSI影响在0.05水平上显著;胸径分段主效应、周向相对位置和胸径分段的二元交互效应接近在0.05水平上显著;其余效应对GSI影响均不显著。进一步对多因素方差分析结果进行分析认为,由于所选取部分倾斜和弯曲的马尾松样木在树干内部可能形成了应压木,所以使轴向生长应力在周向上分布不均,从而导致周向相对位置对GSI产生了显著影响。另外可以看出,树高分段和胸径分段对GSI存在一定的影响,虽然这种影响没有达到统计学显著水平,但可以推断,马尾松立木的树高和胸径在一定程度对立木轴向表面生长应力存在影响,2个性状可以用来对生长应力进行早期评估,并以此为参照标准来进行生长应力调控;而2个冠形指标,是否偏冠和冠幅大小无论是主效应还是交互效应对GSI的影响都不显著,这说明不能通过是否偏冠和冠幅大小来调控和评估人工林马尾松轴向生长应力大小。
表 4中3个定性自变量的两两比较结果显示,3种不同树干形态可分为2个子集:(直立、倾斜)、(倾斜、弯曲),差异在0.05水平显著,平均值大小依次为:直立>倾斜>弯曲;8个不同周向相对位置可以分为3个子集,3个子集之间差异都在0.05水平显著,3个子集分别为:(1、2、5、8、4、7、)、(2、5、8、4、7、3)、(6),其平均值大小依次为1>2>5>8>4>7>3>6。产生上述比较结果的原因同样可能是由于部分倾斜和弯曲样木中在倾斜下部或弯曲凸处(标示为1位置)产生了应压木,导致表面轴向生长应力由拉应力转变为压应力,GSI数值也随之变为负值,由于直立树干在周向上不可能产生轴向压应力,所以均值大于其他2种树干形态。
采集地两两比较将3个不同采集地分为2个子集,即沙河集一个子集,八公山纯林和混交林为一子集,差异性显著性达到0.05水平。各采集地GSI平均值大小顺序为:沙河集>八公山混(八公山混交林简称,下同)>八公山纯(八公山纯林简称,下同)。通过平均值可以明显看出不同采集地区之间差异较大,但同一采集地区不同林型之间几乎没有差异,这说明马尾松通过与麻栎混交产生的复层林无法对轴向表面生长应力产生显著影响,这一结果与Saurant等(1976)对欧洲水青冈研究得出的复层林能有效降低生长应力结果不一致。从2块采集地马尾松倾斜和弯曲样木生长性状数据和弯曲树干草图比较可以看出,八公山林场样木倾斜树干倾斜角度要大于沙河集林场,弯曲树干测试区域的弯曲程度也同样大于沙河集林场马尾松样木,所以在倾斜树干和弯曲树干中产生应压木的机率也应大于沙河集林场马尾松样木,因此八公山林场GSI均值在比较中处于劣势。对3个定性变量的两两比较结果进一步分析,则可以认为马尾松产生应压木可能性的大小关系到不同树干形态、不同周向相对位置乃至不同采集地对GSI的影响程度,因此可以通过对应压木产生可能性的大小的估测来对GSI的大小和分布进行预测。
通过对不同树干形态马尾松GSI的比较研究,可以看出马尾松倾斜树干和弯曲树干可能存在应压木,在不同周向相对位置中轴向表面生长应力差异极显著,所以立木轴向表面生长应力可以对马尾松产生应压木的几率和它所占比例进行预测。直立树干由于产生应压木几率很小,所以周向上生长应力差异不显著,而且其均值也大于倾斜和弯曲树干。
2.2 不同树干形态马尾松生长性状与GSI之间的关系利用SPSS软件中数据转换过程将几个定性描述变量转变成序数变量,如用1、2分别表示树冠大小的窄、阔和是否偏冠的正冠、偏冠,不同周向位置直接将标示号转变成序数。为准确说明某一指标对轴向表面生长应力直接影响的大小和得出各指标对轴向表面生长应力直接影响程度的大小次序,本文引入偏相关分析来研究3种不同树干形态中各自变量与GSI的关系,在计算任一自变量与GSI之间的偏相关系数时,控制其余所有变量,所得结果如表 5所示。
|
|
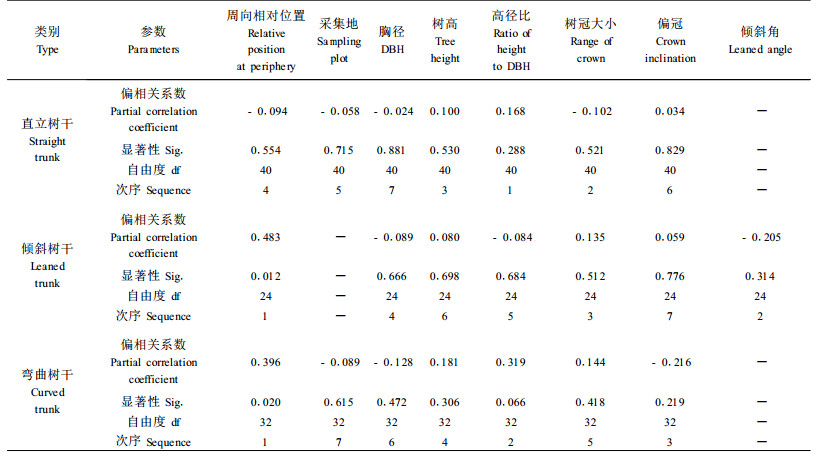
由表 5可知,在直立树干中,所有自变量与GSI之间的偏相关系数均不显著,但是偏相关系数的绝对值可以在一定程度上反映该自变量对GSI的直接影响程度,可以用它作为标准来衡量各自变量对GSI影响程度的相对大小。直立树干各自变量对GSI影响程度大小次序为:高径比>树冠大小>树高>周向相对位置>采集地>胸径>是否偏冠。从表 5中可以观察到,GSI与高径比呈正偏相关,与树高呈正偏相关,与胸径呈负偏相关,这说明高径比越大,轴向表面生长应力越大;另外在多因素方差分析中也得出了树高和胸径能在一定程度影响轴向表面生长应力,因此高径比可以用来作为对轴向表面生长应力进行预测和调控的参照指标,并且通过合理地间伐来降低立木高径比可以在一定程度上降低生长应力,这与Polge(1981)研究欧洲水青冈得出的结论相类似。2个冠形变量中,直立树干GSI与树冠大小之间为负偏相关,与是否偏冠之间为正相关,但2个偏相关关系均不显著,通过结合变量转化过程可知,直立马尾松立木的偏窄树冠轴向表面生长应力较大。另外本文中树冠大小在偏相关系数绝对值排序中为第二位,所以可以认为树冠的大小对直立树干GSI存在一定的直接影响,可以作为参照生长性状指标对马尾松直立立木的轴向表面生长应力进行预测和调控,而是否偏冠次序和采集地次序靠后不能作为参照指标。
2.2.2 倾斜树干生长性状与轴向GSI之间的关系在倾斜树干中,周向相对位置与GSI之间正偏相关系数在0.05水平上显著,其余自变量与GSI之间偏相关系数均不显著。同样利用偏相关系数的绝对值来比较倾斜树干各自变量对GSI直接影响程度大小,其结果为:周向相对位置>倾斜角>树冠大小>胸径>高径比>树高>是否偏冠。周向相对位置编号顺序是从倾斜树干的下部向上部逐渐增大,所以GSI与周向相对位置的正偏相关关系表示轴向表面生长应力从倾斜树干下部到上部逐渐增大,且这种关系在0.05水平上显著。很明显这是由于部分马尾松树干在倾斜下部产生应压木,轴向表面应力由拉应力转变成压应力,从而在此处GSI产生负值,形成这种显著的正偏相关关系。利用勾股定理计算出来的倾斜树干倾斜角与GSI之间为负偏相关关系,这说明倾斜角越大的马尾松立木,其轴向表面应力均值越小。产生这种关系的原因也是由于应压木的形成,而且由于树干倾斜角度越大,应压木产生的几率和在树干中的比例也应随着增大,所以在周向上出现GSI为负值的测试点和测试点负值的绝对值也相应增加,导致了随着倾斜角的增大,周向上GSI均值开始下降,这个研究结果与本文多因素方差分析中3个定性变量两两分析结果所得的推论较为一致,也进一步证实了马尾松立木中存在应压木的几率和应压木区域所占比例对立木表面轴向生长应力有一定的影响。倾斜树干树高、胸径和高径比3个生长性状与GSI值之间的偏相关关系较弱,并且GSI与树高之间的正相关、胸径的负相关与GSI与高径比之间的负相关出现矛盾,原因可能是由于本身偏相关系数数值较小,另外还控制了其余变量,所以造成了结果之间的偏差,因此可以认为倾斜树干的树高和胸径对轴向表面生长应力直接影响很小,不能作为倾斜树干轴向表面生长应力评估和控制的参照指标。在分析冠性变量与GSI的偏相关系数中发现,具有阔偏树冠的倾斜树干轴向表面生长应力更大,同样由于树冠大小在绝对值排序中位置靠前,所以在本文中认为可以将它列为参照指标,而是否偏冠仍然是排序较后,所以不能作为参照指标。
2.2.3 弯曲树干生长性状与轴向GSI之间的关系在弯曲树干研究中,周向相对位置与GSI之间正偏相关系数仍然在0.05水平上显著,另外,高径比与GSI之间正偏相关系数接近在0.05水平上显著。弯曲树干各自变量对GSI直接影响程度大小为:周向相对位置>高径比>是否偏冠>树高>树冠大小>胸径>采集地。周向相对位置与GSI之间正偏相关系数在0.05水平上显著的原因和倾斜树干类似,不同的是弯曲树干的GSI是从弯曲凸处向凹处逐渐增大。根据弯曲树干的高径比与GSI之间偏相关关系得出的推论与在直立树干中得出的推论相同,即可以将高径比作为参照指标,并且树高和胸径与GSI之间的偏相关关系也证明了这一点。弯曲树干的2个冠性变量与GSI之间的偏相关关系反映出具有窄正树冠的马尾松轴向表面生长应力较大,是否偏冠指标对GSI的直接影响在排序中为第三,所以本文中将它列为马尾松弯曲树干轴向表面生长应力的参照指标,而树冠大小排序较后不作为参照指标,采集地排序中最末,也不能作为参照指标。
3 结论与讨论1) 多因素方差分析得出,主效应周向相对位置对GSI影响在0.001水平上显著;周向相对位置和树高分段的二元交互效应对GSI影响在0.05水平上显著,其余效应对GSI影响均不显著。S-N-K法两两比较将3个树干形态和3个采集地分成2个子集,将8个不同周向相对位置分成3个子集。
2) 偏相关分析研究中,在直立树干所有自变量与GSI之间的偏相关系数均不显著;倾斜树干中,周向相对位置与GSI之间正偏相关系数在0.05水平上显著,其余自变量与GSI之间偏相关系数均不显著;弯曲树干中,周向相对位置与GSI之间正偏相关系数也在0.05水平上显著。直立树干中,GSI与高径比、树高、是否偏冠之间成正偏相关关系,与胸径、树冠大小成负偏相关;倾斜树干中,GSI与倾斜角、胸径、高径比之间为负偏相关关系,与树高、是否偏冠、树冠大小之间为正偏相关;弯曲树干中,GSI与高径比、树高、树冠大小之间为正偏相关,与其余因子呈负偏相关关系。
3) 综合分析多因素方差分析和偏相关分析表明,对于不同树干形态,立木的高径比、倾斜角、冠形因子对表面轴向生长应力的影响程度不尽相同。本文认为可以通过间伐降低直立树干和弯曲树干的高径比,从而能在一定程度上降低马尾松立木的轴向表面生长应力。通过控制倾斜角和冠性因子也可以一定程度上影响倾斜树干轴向表面生长应力的大小和分布。
安徽农学院林学系. 1980. 马尾松. 北京: 中国林业出版社.
|
胡继青, 姜笑梅, 侯祝强, 等. 2000. 三种人工林桉树轴向生长应变变异初探. 木材工业, 14(6): 9-11. |
黄振英. 2004. 马尾松正常木与应压木生长应力及材性的比较研究. 安徽农业大学硕士学位论文. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y607264
|
费本华, 江泽慧, 赵荣军, 等. 2004. 桉树人工林木材生长应变研究. 木材工业, 18(2): 18-20. DOI:10.3969/j.issn.1001-8654.2004.02.005 |
林金星, 李正理. 1993. 马尾松正常木与应压木的比较解剖. 植物学报:英文版, 35(3): 201-205. |
Archer R R. 1987. Growth stresses and strains in trees. Berlin:Springer-Verlag: New York, 1-5.
|
Bamber R K. 1979. The origin of growth stresses in trees. Part Ⅰ. Micromechanics of the developing cambial cell wall. Wood Sci Technol, 21: 139-154. |
Bamber R K. 1987. The origin of growth stresses in trees: a rebuttal. IAWA Bull, 8: 80-84. DOI:10.1163/22941932-90001032 |
Boyd J D. 1972. Tree growth stresses. Part Ⅴ. Evidence of an origin in differentiation and lignification. Wood Sci and Technol, 6: 251-262. DOI:10.1007/BF00357047 |
Chafe S C. 1979. Growth stress in trees. Australia Forestry Research, 9: 203-223. |
Clair B, Ruelle J, Thibaut B. 2003. Relationship between growth stresses, mechano-physical properties and proportion of fibres with gelatinous layer in chestnut(Castanea sativa Mill.). Holzforschung, 57: 189-195. |
Ferrand J C. 1982. Growth stresses and silviculture of Eucalyptus. Australia Forest Research, 13: 75-81. |
Fournier M, Chanson B, Thibaut B, et al. 1994. Mesure des déformations résiduelles de croissance à la surface des arbres, en relation avec leur morphologie. Observation sur différentes espèces. Observation sur différentes espèces.Ann Sci For, 51: 249-266. |
Gerard J, Bailleres H, Fournier M, et al. 1995. Wood quality in plantation Eucalyptus:a study of variation in three reference properties. Bois et Forêts des Tropiques, 245: 101-110. |
Malan F S, Gerischer G F R. 1987. Wood property differences in South African grown Eucalyptus grandis trees of different growth stress intensity. Holzforschung, 41(6): 331-335. DOI:10.1515/hfsg.1987.41.6.331 |
Okuyama T, Kangagawa Y, Hattori Y. 1987. Reduction of residual stresses in logs by direct heating method. Mokuzai Gakkaishi, 33(11): 837-843. |
Okuyama T, Sasaki Y, Kikata Y, et al. 1981. The seasonal change in growth stress in the tree trunk. Mokuzai Gakkaishi, 27(5): 350-355. |
Polge H. 1981. Influence des eclaircies sur les contraintes de croissance du hêtre. Annal of Forest Science, 38: 407-423. DOI:10.1051/forest:19810401 |
Saurat J, Gueneau P. 1976. Growth stresses in beech. Wood Science and Technology, 111-123.
|
Wang G. 1972. Growth stresses-genetic and environmental influences. News letter 389. FPL CSIRO Melbourne For Conversion Eng Group: 3-4.
|
Wang G. 1977. Reducing growth stresses in standing trees. Australia Forest Research, 7: 215-218. |
Watanabe H. 1965. A study of the origin of longitudinal growth stresses in tree stems. Proc IUFRO Sect 41, Melbourne, Aust Committee on growth stresses.
|
Yamamoto H. 1998. Generation mechanism of growth stresses in wood cell walls: Roles of lignin deposition and cellulose microfibril during cell wall maturation. Wood Science and Technology, 32: 171-182. DOI:10.1007/BF00704840 |
Yang J L, Gary W. 2002. Growth stress, it measurement and effects. Australia Forestry, 64(2): 127-135. |