 2008, Vol. 44
2008, Vol. 44文章信息
- 金国庆, 秦国峰, 刘伟宏, 储德裕, 洪苏州, 周志春.
- Jin Guoqing, Qin Guofeng, Liu Weihong, Chu Deyu, Hong Suzhou, Zhou Zhichun.
- 马尾松生长性状交配效应的遗传分析及杂交组合选择
- Effects of Mating Manners on Growth Traits of Pinus massoniana and Selection of Cross Combinations
- 林业科学, 2008, 44(6): 28-33.
- Scientia Silvae Sinicae, 2008, 44(6): 28-33.
-
文章历史
- 收稿日期:2007-05-10
-
作者相关文章


2. 浙江省淳安县姥山林场 淳安 311700
2. Laoshan Forest Farm of Chun'an County, Zhejiang Province Chun'an 311700
杂交育种是植物遗传改良有效的育种手段和主要途径之一,包括亲本选配、子代测定和新品种(系)鉴定三方面工作(李周岐等,2001)。其中,亲本选配是杂交育种成败的先决条件(张爱民,1994);子代测定可估算亲本无性系育种值,用于后向选择,作为对原建种子园无性系留优去劣的依据,用于前向选择,在全同胞优良家系中优选个体为高世代遗传改良提供育种材料(王明庥,2001)。此外配合力育种还可了解树种重要性状的基因作用方式,制订有效的遗传改良方案等(齐明,1996)。开展不同遗传交配设计的杂交育种,旨在根据某一树种不同的培育目标,创制高产优质高抗新品种。双列交配设计可以同时估算一般配合力(GCA)和特殊配合力(SCA)等遗传参数,而且具有统计上的稳定性,是一种有效的参数估算方法(Kearsey,1965;Pederson,1972),因而在作物和林木遗传育种中得到广泛应用(Huber et al., 1991;King et al., 1997;李淡清等,2002;李开隆等,2006)。火炬松(Pinus taeda)(Sluder,1993)、湿地松(P.elliottii)(Sluder,1996)、辐射松(P.radiata)(Wilcox,1982)及杉木(Cunninghamia lanceolata)等重要针叶用材树种都曾开展过双列交配设计和配合力研究(叶培忠等,1981;王赵民等,1988;叶志宏等,1991;支济伟等,1994;李力等,2000)。
马尾松(Pinus massoniana)是我国南方具有巨大经济价值和生态效益的主要乡土造林树种,其花粉是高级天然营养保健食品及美容养颜珍品,松脂是生产松香和松节油的原料,木材是重要的造纸等工业用材。该树种自然分布范围广、地理种源变异丰富、适应性强,具有巨大的遗传改良前景。因此,该树种自“六五”以来,一直被列为国家重点科技攻关树种之一,先后开展了种源试验、种子园营建技术及杂交育种研究等。对马尾松开展双亲控制授粉的杂交育种研究起步较晚,目前只有周志春等(2004)对马尾松纸浆材重要经济性状配合力及杂种优势进行了研究。本文利用设置在浙江省淳安县姥山林场的2片完全双列交配设计的子代遗传测定林材料,开展亲本的配合力、自交和异交效应分析,以揭示性状的遗传控制方式,评价杂交亲本和组合,研究自交衰退和种内杂种优势等问题,为马尾松高世代育种和种内杂种优势利用提供理论依据。
1 材料与方法 1.1 试验材料浙江省淳安县姥山林场马尾松育种园共收集来自南方11个省区1 000多个优树无性系。1986—1988年嫁接,1992年起有大量的无性系开花结实。分别于1992年和1993年连续2年开展6×6完全双列交配设计杂交制种,其亲本无性系完全相同,分别来自广东(1003)、江西(6610、6627)、浙江(5163、5907)、安徽(3412)等产地。随后3年分别育苗,并在姥山林场营造子代测定林。造林试验采用完全随机区组设计,8株小区,5次重复,试验林位于119°03′E,29°37′N,海拔250~300 m,年平均气温17 ℃,≥10 ℃的年积温为5 410 ℃,年降水量1 430 mm,年日照1 951 h。在育苗与造林时,用浙江省淳安县当地天然优良采种林分子代作对照(CK),2片试验林在11年生(材料Ⅰ)与10年生(材料Ⅱ)时进行每木调查,测定树高和胸径等性状值。
1.2 统计分析根据测定的树高、胸径与按公式V = 0.000 062 341 803×D1.31.855 149 7×H0.956 824 92估算的单株材积数据进行统计分析。以试验小区平均值为单位,采用SAS软件包中的GLM程序进行方差分析以检验不同杂交组合间的遗传差异显著性。若杂交组合效应显著,按Griffing双列杂交方法Ⅰ,以组合平均数进行配合力方差分析,按模型Ⅰ(固定模型)估算一般配合力(GCA)、特殊配合力(SCA)和正反交效应值(REC),按模型Ⅱ(随机模型)估算遗传方差分量等(Griffing,1956;马育华,1982;王明庥,1989;朱军,1997;续九如,2006),性状的配合力方差分析模型列于表 1。自交衰退程度(Di)的估算按Kheradnam等(1975)方法,其公式为:Di =(FSi-OP)/OP×100%,式中:Di为自交衰退程度,FSi为任一亲本自交F1代家系的平均值,OP为(任)某一自交家系相应亲本的自由授粉子代平均值。杂交组合的杂种优势评定用类似Bahman等(1975)方法,其公式为:H=(Fi-OP)/OP×100%,式中:H为杂种优势程度,Fi为任一个杂交组合(家系)的平均值,OP为(任)某一个Fi组合中一个优势亲本的自由授粉子代的平均值。
|
|

方差分析(表 2)表明,所测试的2份材料中除材料Ⅱ的胸径与树高在父本间的遗传差异达5%的显著水平外,其余树高、胸径和材积各性状在组合间、母本间和父本间均达到了1%的极显著差异水平。这说明通过马尾松种内不同种源(或产地)的无性系间杂交,可为下一世代的育种创造变异丰富的基本群体,使创制和筛选优良杂交组合成为可能。
|
|
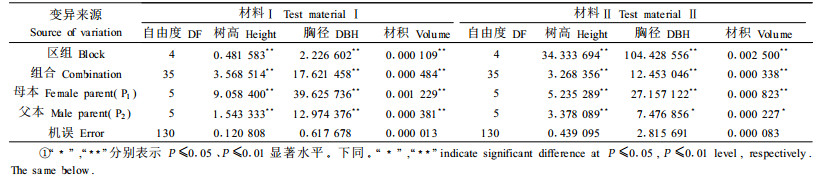
进行性状配合力分析,揭示其基因作用方式,可正确有效地制定育种方案。配合力方差分析结果(表 3)表明,除材料Ⅱ中树高的正反交效应达5%显著水平外,2份材料中其余树高、胸径和材积各性状的一般配合力(GCA)、特殊配合力(SCA)和正反交效应(REC)均达到了1%的极显著水平。
|
|
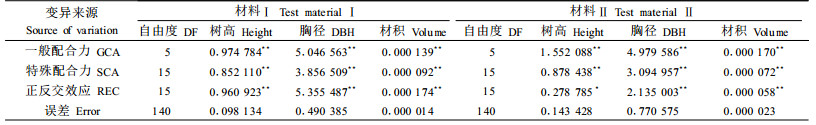
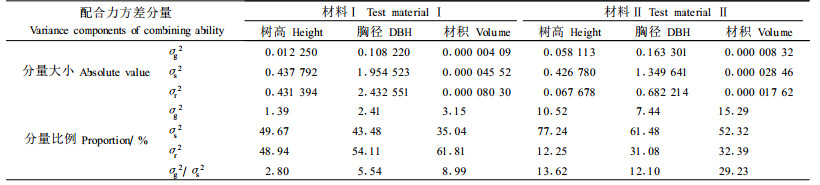
根据全同胞的遗传方差理论,GCA、SCA和REC方差主要是由加性基因效应、显性效应和上位效应引起的(Mullin et al., 1992),因此,可根据配合力方差分量大小,来说明性状遗传控制方式。从表 4结果来看,马尾松主要生长性状的一般配合力方差远小于特殊配合力方差,2份材料中树高、胸径和材积各性状的σg2/σs2分别为2.80%~13.62%、5.54%~12.10%和8.99%~29.23%,说明显性基因效应起主要作用,尤其在材料Ⅱ中居主导地位,而加性基因效应相对较小。各生长性状的正反交效应方差分量所占比例因试验材料不同而有较大差别,在材料Ⅰ中树高、胸径和材积的值分别达48.94%、54.11%和61.81%,表明上位效应起主导作用,在材料Ⅱ中树高、胸径和材积的值分别为12.25%、31.08%和32.39%,说明上位效应起较大作用。
|
|
6个亲本自交效应分析结果(表 5)表明,马尾松各亲本自交后代在各生长性状上普遍存在衰退现象,其自交衰退程度(Di)依测试材料和杂交亲本而异。在材料Ⅰ中,各亲本树高、胸径和材积的Di分别为-8.0%(1003)~-28.7%(6610)、-12.5%(1003)~-40.5%(6610)和-16.8%(1003)~-71.6%(6610),平均为-17.8%、-27.7%和-50.6%。在材料Ⅱ中,各亲本树高、胸径和材积的Di分别为-4.6%(6627)~-45.3%(6610)、-6.8%(1003)~-50.8%(6610)和-14.5%(1003)~-84.5%(6610),平均为-18.4%、-23.3%和-44.3%。在2份材料中,亲本6610的自交衰退程度最高,而亲本1003的自交衰退程度最小。
|
|

通过性状的Di与相应的GCA相关分析结果显示,2份材料中亲本自交衰退程度与其一般配合力大小在树高、胸径和材积各性状中均存在显著或极显著的相关性,说明一般配合力小的亲本,其自交衰退程度会有明显增加的趋势。
不同研究材料各杂交组合与优亲自由授粉子代之间存在着不同的优势效应。从表 6列出的11年生和10年生2片马尾松全同胞子代测定林主要生长性状的杂种优势分析结果发现,不同杂交组合间的杂种优势差异很大。在材料Ⅰ中,树高、胸径和材积各生长性状的超亲优势在杂交组合间变化分别为-19.15%~ 18.57%、-24.14~35.36%和-50.44%~82.48%,其中6627×6610、1003×3412、6627×3412、6627×1003、5163×3412、6627×5907、1003×6610、5163×6627、3412×1003、1003×5907、5163×1003这11个组合具有较强的杂种优势,其材积杂种优势都在20%以上,而5907×6627、3412×6627、6610×1003、1003×6627等的材积则呈现明显的杂种劣势。在材料Ⅱ中,树高、胸径和材积各生长性状的超亲优势在杂交组合间变化分别为-14.02%~11.27%、-18.40~26.07%和-36.43%~46.63%,其中6627×5163、1003×3412、6627×6610、6627×1003、6610×5163、1003×5907这6个组合具有较强的杂种优势,其材积杂种优势均在15%以上,而3412×6627、5907×1003、5163×6610、3412×5163等的材积则呈现明显的杂种劣势。比较分析发现,相同亲本组合的正反交杂种优势有很大差异甚至出现相反的结果。从总体上看,2份材料大致呈相同的规律。
|
|
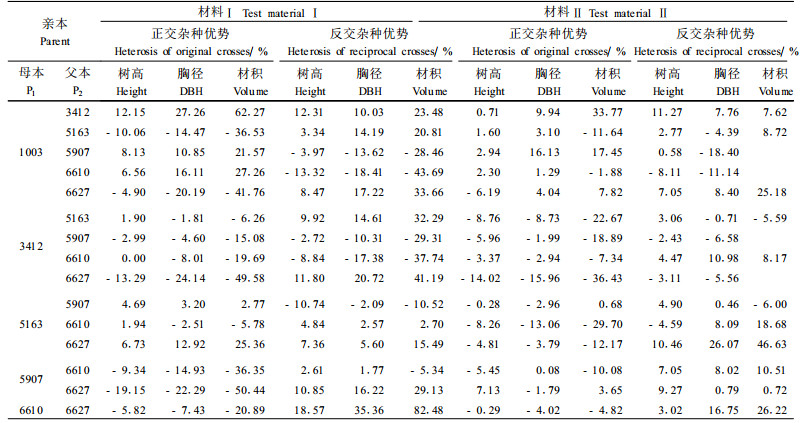
在分析的3个生长性状中,与经济价值关系最为密切的是材积。在不同杂交组合杂种优势分析的基础上,以材积生长量作为优良杂交组合评选的主要指标。材料Ⅰ中选出1003×3412、6627×6610、6627×1003、1003×6610、3412×1003、1003×5907、5163×1003、5163×3412、6627×3412、5163×6627共10个优良杂交组合。如以试验平均为对照,优良杂交组合的材积现实遗传增益为38.17%~101.98%;若以CK为对照,优良杂交组合的材积现实遗传增益高达73.86%~154.16%。同样,材料Ⅱ中选出6627×5163、1003×3412、6627×1003、1003×5907、6627×6610、5163×1003、1003×6627、3412×1003、6610×5163、1003×6610共10个优良组合。如以试验平均为对照,优良杂交组合的材积现实遗传增益为20.80%~67.92%;若以CK为对照,优良杂交组合的材积现实遗传增益则达48.80%~106.84%。2份材料中所选出的各10个优良杂交组合中有7个组合是相同的,详见表 7。
|
|

双亲材料的配合力测定是杂交育种工作的重要内容,估算GCA、SCA和REC的大小及其相对重要性,可揭示性状的遗传控制模式,科学地制定有效的育种策略。配合力的大小和相对重要性与测定材料有关。本研究中观察到马尾松主要生长性状具显著的GCA、SCA和REC效应,说明这些性状的遗传是由加性和非加性基因共同控制,其中非加性效应(显性效应和上位效应)起主导地位,加性基因效应相对较小。虽然2种配合力的相对重要性存在较大差异,但是,无论GCA与SCA方差分量大小如何,只要其相应的配合力效应显著,对其进行测定与选择都是非常必要的。基因的上位效应因测定材料而异,材料Ⅰ中明显大于材料Ⅱ中,这主要是因为2片试验林的立地条件差异较大,试验中存在明显的基因与环境(G×E)互作效应所致。本研究得到的马尾松GCA和SCA的相对重要性与玉米(Zea mays)和杉木的研究结果不同(齐明,1996;李力等,2000;周志春等,2004),这可能有两方面的原因:其一,本试验所选的6个交配亲本虽未经过生长性状的一般配合力的测定,但由于当初在选配杂交亲本时,都选择一些开花年龄较早的无性系,从而在开花习性上对6个交配亲本生长性状的一般配合力进行了间接选择;其二,本试验所用交配亲本来源于不同的地理种源区,根据杂交育种的理论,遗传距离较大的优良无性系进行杂交可望产生特殊配合力较强的杂交组合,在全部36个杂交组合中,相当多的组合代表了不同种源间杂交。因此,对马尾松的遗传改良通过建立种子园利用GCA效应的同时,还应通过创制高特殊配合力的杂交组合,走有性制种无性规模扩繁利用的路径。
马尾松是自交可育性较强的树种,在种子园投产初期自交率可高达30%左右(金国庆等,2000)。马尾松各亲本无性系的自交后代在树高、胸径和材积各生长性状上普遍存在衰退现象,其自交衰退程度随测试材料和杂交亲本无性系而异。在2份材料中,各亲本树高、胸径和材积的平均自交衰退程度分别达-17.8%~-18.4%、-23.3%~-27.7%和-44.3%~-50.6%。同时还发现,各生长性状的自交衰退程度与其一般配合力大小呈显著或极显著的相关性,表明一般配合力小的亲本,其自交衰退程度有明显增加的趋势。
强优势杂交组合的创制首先是杂交亲本的选配。王赵民等(1988)通过对杉木生长性状的配合力和杂种优势研究认为,利用一般配合力高的亲本进行交配将会提高得到杂种优势的机率。本研究发现,马尾松不同交配组合间的杂种优势差异巨大,如6627×6610和1003×3412等组合的生长性状具有较强的杂种优势,而3412×6627等组合则表现出明显的杂种劣势。此外,研究还发现,一般配合力较高亲本的自交衰退程度较低,自交不衰退或衰退程度较低可能与2个亲本具有较高的一般配合力效应有关,这一现象在杉木等树种中也有发现(陈岳武等,1982)。在优良组合的评选中,由于存在着显著的正反交效应,在这种情况下,根据特殊配合力效应值选择杂交组合时应谨慎,应在不同杂交组合杂种优势分析的基础上,以材积生长量作为优良杂交组合评选的主要指标,参考SCA及GCA等进行优良杂交组合选择,这样选出的优良杂交组合具有较强的杂种优势。生产上为了充分利用其杂种优势,应对优良杂交组合中的优株进行无性繁殖。另外,由于马尾松生长性状存在明显的自交衰退和正反交效应,在营建杂交种子园,尤其是双系杂交种子园时,依据特殊配合力效应值选择与配置亲本时,应考虑到亲本无性系的自交效应及反交效应等影响,避免这些情况的发生。
陈岳武, 施季森, 刘大林. 1982. 杉木种内杂种优势及亲本配合力的分析. 南京林产工业学院学报, (2): 1-18. |
金国庆, 秦国峰, 周志春, 等. 2000. 马尾松无性系种子园结实初期异交水平分析. 林业科学研究, 13(5): 464-468. DOI:10.3321/j.issn:1001-1498.2000.05.002 |
李淡清, 刘永平, 曾德贤, 等. 2002. 蓝桉6×6全双列交配生长性状的遗传效应分析. 遗传学报, 29(9): 835-840. |
李开隆, 姜静, 姜莹, 等. 2006. 白桦5×5完全双列杂交种苗性状的遗传效应分析. 北京林业大学学报, 28(4): 82-87. DOI:10.3321/j.issn:1000-1522.2006.04.016 |
李力, 施季森, 陈孝丑, 等. 2000. 杉木两水平双列杂交亲本配合力分析. 南京林业大学学报, 24(5): 9-13. DOI:10.3969/j.issn.1000-2006.2000.05.003 |
李周岐, 王章荣. 2001. 鹅掌楸属种间杂种苗期生长性状的亲本配合力分析. 西北林学院学报, 16(3): 7-10. DOI:10.3969/j.issn.1001-7461.2001.03.002 |
马育华. 1982. 植物育种的数量遗传学基础. 南京: 江苏科学技术出版社.
|
齐明. 1996. 杉木育种中GCA与SCA的相对重要性. 林业科学研究, 9(5): 498-503. DOI:10.3321/j.issn:1001-1498.1996.05.010 |
王明庥. 1989. 林木育种学概论. 北京: 中国林业出版社, 230-236.
|
王明庥. 2001. 林木遗传育种学. 北京: 中国林业出版社, 1-5.
|
王赵民, 陈益泰. 1988. 杉木主要生长性状配合力分析及杂种优势的利用. 林业科学研究, 1(6): 614-620. |
续九如. 2006. 林木数量遗传学. 北京: 高等教育出版社.
|
叶培忠, 陈岳武, 刘大林, 等. 1981. 配合力分析在杉木数量遗传学中的应用. 南京林产工业学院学报, (3): 1-21. |
叶志宏, 施季森, 翁玉秦, 等. 1991. 杉木11个亲本双列交配遗传分析. 林业科学研究, 4(4): 380-385. |
张爱民. 1994. 植物育种亲本选配的理论和方法. 北京: 中国农业出版社, 1-12.
|
支济伟, 陈益泰, 骆秀琴, 等. 1994. 杉木主要材质性状配合力研究. 林业科学研究, 7(5): 531-536. DOI:10.3321/j.issn:1001-1498.1994.05.013 |
周志春, 金国庆, 秦国峰, 等. 2004. 马尾松纸浆材重要经济性状配合力及杂种优势分析. 林业科学, 40(4): 52-57. DOI:10.3321/j.issn:1001-7488.2004.04.009 |
朱军. 1997. 遗传模型分析方法. 北京: 中国农业出版社.
|
Bahman Y S, Sarafi A, Zali A A. 1975. Heterosis and inbreeding estimates in Safflower. Crop Science, 15(1/2): 81-83. |
Griffing B. 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust J Biol Sci, (9): 463-493. |
Huber D A, White T L, Littell R C, et al. 1991. Ordinary least squares estimation of general and specific combining abilities from half-diallel mating designs. Silv Genet, 41(4/5): 263-273. |
Kearsey M J. 1965. Biometrica1 analysis of a random mating population:a comparison of five experimental designs. Heredity, 20(3): 205-235. |
Kheradnam M, Bassire A, Niknejad M. 1975. Heterosis, inbreeding depression, and reciprocal effects for yield and some yield components in a Cowpea cross. Crop Science, 15(9/10): 689-691. |
King J N, Carson M J, Johnson G R. 1997. Analysis of disconnected diallel mating designs Ⅱ: Results from a third generation progeny test of the new Zealand radiata pine improvement programme. Silv Genet, 47(2/3): 80-87. |
Mullin T J, Park Y S. 1992. Estimating genetic gains from alternative breeding strategies for clonal forestry. Can J For Res, 22(1): 14-23. DOI:10.1139/x92-003 |
Pederson D G. 1972. A comparison of four experimental designs for the estimation of heritability. Theor Appl Genet, 42(3): 371-377. |
Sluder E R. 1993. Results at age 15 years from a half-diallel cross among 10 loblolly pines selected for resistance to fusiform rust (Cronartium querctmm f.sp.fusiforme). Silvae Genetica, 42(4/5): 223-230. |
Sluder E R. 1996. Two-stage selection in slash pine produces good gains in fusiform rust resistance. Southern Journal of Applied Forestry, 20(3): 143-147. |
Wilcox M D. 1982. Genetic variation and inheritance of resistance to Dothistroma needle blight in Pinus radiata. New Zealand Journal of Forestry Science, 12(1): 14-35. |