 2008, Vol. 44
2008, Vol. 44文章信息
- 王兴昌, 王传宽, 张全智, 李世业, 李国江.
- Wang Xingchang, Wang Chuankuan, Zhang Quanzhi, Li Shiye, Li Guojiang.
- 东北主要树种心材与边材的生长特征
- Growth Characteristics of Heartwood and Sapwood of the Major Tree Species in Northeastern China
- 林业科学, 2008, 44(5): 102-108.
- Scientia Silvae Sinicae, 2008, 44(5): 102-108.
-
文章历史
- 收稿日期:2007-03-05
-
作者相关文章



大多数树木的树干包括边材和心材2部分。边材是指新近几年由维管形成层向内分裂、生长及分化而形成的次生木质部,包括具有输导液流和支持作用的导管和管胞、具有贮藏养分作用的薄壁细胞。在边材的里面即心材,是由死细胞组成的色泽较深的中央锥体,其细胞失去生理活性,并沉积了大量树脂、胶质、鞣质、油类和色素等物质。边材向心材的转化过程是树木生长中一个重要的生理学过程。早在1964年提出的“管道模型"理论就阐明了树木叶量或叶面积与树干的边材面积呈正相关关系(Shinozaki et al., 1964)。边材面积与叶面积之比也常常用于表征树木的蒸腾耗水或固碳能力(Makela et al., 2001)。此外,心材和边材材性上的差异也会影响木材的加工和利用(Longuetaud et al., 2006)。例如,欧洲赤松(Pinus sylvestris)作为用材林时其坚固耐用的心材更为人们青睐(Bjorklund,1999);而作为纸浆材,人们则更倾向于利用含次生物质较少的边材。因此,研究树木边材和心材生长发育规律,是探索树木水分关系和碳代谢机理、构建树木生长收获模型、有效利用木材的基础。
对某一株树木而言,其边材量(宽度)是相对稳定的,但其心材量(半径)则随树木的生长而逐年增加(汪矛等,1998)。然而,不同树种之间,即使同一树种的不同植株之间心材和边材量都存在一定的差异,主要受树木生长(Pinto et al., 2004;Knapic et al., 2005;Miranda et al., 2006;Wang et al., 2007)、立地条件(Climent et al., 2002;Bektas et al., 2003)、森林培育措施(Wilkins,1991;Morling et al., 1999;Miranda et al., 2006)、测树学参数(Wilkes,1991;Climent et al., 2002;2003)、遗传特性(Ericsson et al., 1999;Woeste,2002)等因素的影响。东北温带森林在我国林业和生态环境中占有重要的地位,但目前对其主要组成树种的心材和边材生长发育及其变化规律尚不太清楚。为此,本文以东北典型温带次生林7个主要组成树种——红松(Pinus koraiensis)、兴安落叶松(Larix gmelinii)、春榆(Ulmus japonica)、水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica)、黄菠萝(Phellodendron amurense)和蒙古栎(Quercus mongolica)为研究对象,比较分析各个树种的心材和边材的生长特征,为科学经营和利用东北森林提供科学依据和基础数据。
1 材料与方法 1.1 试验地概况试验地位于黑龙江省帽儿山森林生态系统国家野外科学观测研究站(45°24′N,127°28′E)。该地区属于长白山山系支脉,张广才岭西北部的余脉。植被属于长白植被区系,是东北东部山区典型的天然次生林区。平均海拔300 m,平均坡度10°~15°。土壤多为暗棕色森林土。气候属于大陆性温带季风气候,年降水量600~800 mm,雨量集中在7、8月,年平均蒸发量1 093 mm,年平均相对湿度70%;年平均温度2.7 ℃,全年大于或等于5 ℃的积温为2 897 ℃;年平均日照时数1 850 h,年平均风速1.5 m·s-1。
1.2 野外取样和心边材测定在典型温带森林生态系统中,根据各树种的胸径(diameter breast height,DBH)分布状况随机选取红松、兴安落叶松、春榆、水曲柳、胡桃楸、黄菠萝和蒙古栎各10株,其中包括优势木3株、中等木4株、被压木3株。实测胸径并标明北向后伐倒,从树干基部开始到树梢,每隔1 m截取1圆盘。另外在胸高处追加截取1个圆盘。表 1给出了各树种样木的基本情况。有关林分特征和立地状况、样木的选择和野外作业的详细信息,参见Wang (2006)。
|
|
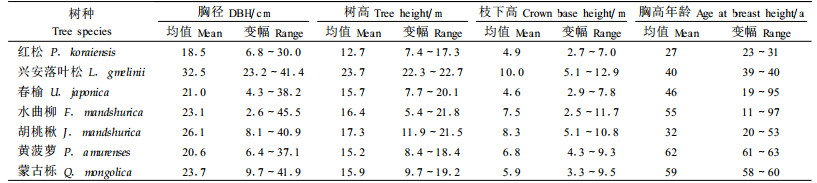
取回的圆盘先将工作面刨光,然后用高精度的专业扫描仪扫描圆盘图像。根据心材与边材颜色差异等区分心边材的界限,并用Windendro2003年轮分析软件(Regent Instruments Co.,Canada)测量每个圆盘东、西、南、北4个方向的总年轮数、边材年轮数(sapwood ring number, SRN)、树干去皮半径(xylem radius, XR)、心材半径(heartwood radius, HR)、边材宽度(sapwood width, SW)。心材年轮数(heartwood ring number, HRN)等于总年轮数与边材年轮数之差。由于部分圆盘的心材和边材难以区分,因此,实际用于分析心材半径和边材宽度的红松、兴安落叶松、春榆、水曲柳、胡桃楸、黄菠萝和蒙古栎的样木数分别为9、6、10、10、10、10和6株;而用于分析心材年轮的样木数分别为9、6、8、9、4、2和0株,分析边材年轮的样木数分别为9、6、8、9、4、7和0株。
1.3 数据分析树干某一高度的圆盘总年轮数表示对应高度的形成层年龄(cambial age, CA)。心材年轮数与形成层年龄回归曲线的斜率表明了心材形成速率(每年边材向心材转化的年轮数);而曲线外推与横轴的交点表明了心材起始树龄(Pinto et al., 2004;Knapic et al., 2005;Longuetaud et al., 2006),即当心材年轮数为0时的形成层年龄(不足0.1按0.1计)。同样,心材半径与树干去皮半径回归曲线的斜率表明了每年边材向心材转化的宽度,曲线外推可得心材起始树干去皮直径(去皮半径的2倍)。
种内和种间边材年轮数和边材宽度差异显著性采用ANOVA和LSD多重比较分析。根据Longuetaud等(2006)的研究,二次模型若没有包含年龄特别小的树木可能会低估心材起始树龄,因此本文采用线性模型估算心材起始树龄。所有的统计分析均采用SPSS 13.0完成。
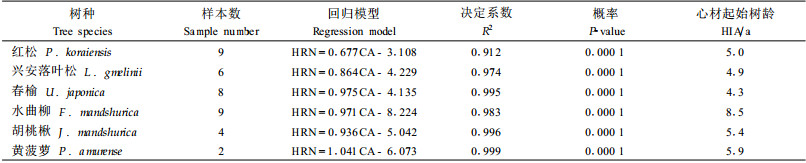
2 结果与分析 2.1 心材生长特征所有树种的心材年轮数都与其形成层年龄呈极显著(P<0.000 1)的正相关关系,形成层年龄能够解释90%以上的心材年轮数变化(表 2)。心材形成速率波动在0.677~1.041 a-1,其中以红松的最小,黄菠萝的最大。被测针叶树心材形成速率均小于阔叶树。心材起始树龄波动在4.3~8.5年,以春榆最小,水曲柳最大。
|
|
各树种的心材半径与树干去皮半径也呈极显著的正相关关系,去皮半径解释了97%以上的心材半径变化(表 3)。各树种心材起始树干去皮直径差异很大,从大到小依次为兴安落叶松、红松、胡桃楸、水曲柳、春榆、蒙古栎和黄菠萝。
|
|

边材年轮数随形成层年龄的变化规律因树种而异(图 1)。红松、春榆、水曲柳和胡桃楸的边材年轮数基本上随着形成层年龄增加而增大;黄菠萝边材年轮数则随着形成层年龄的增加而呈下降趋势;而兴安落叶松边材年轮数开始时随形成层年龄增加而增大,到30年之后则略有下降。

|
图 1 边材年轮数随形成层年龄的变化 Figure 1 Changes in sapwood ring number (SRN) with cambial age (CA) |
与心材半径类似,边材宽度与树干去皮直径也存在显著的正线性相关关系(P<0.000 1),但其决定系数较小,波动在0.124~0.626之间(数据未给出)。
被测树种内植株间边材年轮数和边材宽度差异显著或极显著(春榆边材年轮数例外,P=0.149);树种间的边材年轮数和边材宽度差异均极显著(P<0.001)。LSD多重比较表明:除红松和水曲柳边材年轮数差异不显著、水曲柳与蒙古栎以及胡桃楸与蒙古栎的边材宽度差异不显著外,其余种间差异均为极显著。各树种平均边材年轮数从大到小分别为:红松(9.6±2.8)年、水曲柳(9.5±3.0)年、兴安落叶松(7.9±2.0)年、胡桃楸(6.8±1.7)年、春榆(5.0±1.4)年和黄菠萝(3.9±0.8)年。被测针叶树边材宽度明显大于阔叶树,平均边材宽度从大到小依次为:兴安落叶松(2.55±0.60)cm、红松(1.96±0.72)cm、水曲柳(1.27±0.46)cm、蒙古栎(1.13±0.47)cm、胡桃楸(1.06±0.47)cm、春榆(0.72±0.30)cm和黄菠萝(0.46±0.20)cm。
2.3 心材和边材沿树干的垂直变化所有树种的心材年轮数随高度递减,与总年轮数沿树干的变化格局相同。边材年轮数随高度的变化比心材的变化小得多,但其变化因树种而异(图 2)。红松边材年轮数随高度增大而递减;落叶松边材则先增大后减小;水曲柳却先减小后增大;春榆、胡桃楸和黄菠萝则随高度变化很小。总的来说,针叶树树干上部边材年轮数小于树干中下部,而阔叶树却没有这种格局。

|
图 2 各树种边材年轮数沿树干高度的变化 Figure 2 Changes in sapwood ring number (SRN) along stem height for each tree species |
所有树种心材半径总体上沿树干随高度增大而减小,与树干外形基本一致(图 3)。然而边材宽度随高度变化比心材变化小得多,波动在1.2~2.2 cm之间。几乎所有树木的边材宽度在树干基部异常大,大约从1~2 m开始沿树高的变化不大,平均值波动在0.5 cm之内(图 3)。

|
图 3 7个树种平均木质部半径、心材半径和边材宽度随树高的变化 Figure 3 Changes in mean values of xylem radius (XR), heartwood radius (HR) and sapwood width (SW) with tree height for the seven tree species |
本研究中各个树种的心材年轮数与形成层年龄之间(表 2)、心材半径与树干去皮半径之间(表 3)极显著的正相关关系表明,树木达到一定年龄(心材起始树龄)或直径(心材起始去皮直径)之后,边材就以相对稳定的速率向心材转化(Climent et al., 1993;Sellin,1994;Pinto et al., 2004;Knapic et al., 2006;Miranda et al., 2006)。然而,边材向心材的转化速率和心材的起始树龄或直径却因树种而异(表 2、3),被测针叶树心材形成速率小于阔叶树。本研究的心材形成速率(0.7~1.0 a-1)与以往的研究结果相符(0.5~1.0 a-1)(Hazenberg et al., 1991;Bjorklund,1999;Gjerdrum,2003;Pinto et al., 2004;Knapic et al., 2005)。
与心材生长规律不同,边材年轮数随形成层年龄的变化格局与树种有关。除了黄菠萝之外,其他树种的边材年轮数均随形成层年龄增加而增大(图 1),而边材宽度与树干去皮半径存在着显著的正相关关系。这间接地反映了各个树种的生长规律:随着树木年龄或直径的增长,树冠越来越大,维持整个树体生理活动所需要的输导组织越来越多,边材面积不断增加(Shinozaki et al., 1964)。虽然每年有一定量的边材转换成心材(表 2),但是转换率基本低于1 a-1(黄菠萝例外)。兴安落叶松的边材年轮数在30年后略下降,可能表示此后其生长速率趋向于平稳或下降。然而,黄菠萝则由于其心材形成速率略大于1 a-1(表 2)而使边材年轮数随年龄的增加而略减少(虽然边材年轮数略减,但由于直径生长边材面积仍然增加)。
边材年轮数的树种间差异在一定程度上反映了各树种边材年轮寿命的差异。本研究的边材年轮数波动在3.9~9.6年之间,以红松最高、黄菠萝最低。这表明红松的边材年轮寿命最长、黄菠萝的最短。有报道称日本柳杉(Cryptomeria japonica)边材年轮数可达20~22年(Yang et al., 1994)。
边材是树干中有生理活性的组织,边材断面积是水分输导面积,与叶面积紧密相关(Shinozaki et al., 1964;Wang,2006)。所以,在直径相同时,边材宽度的种间差异表明了种间水分利用对策、光合生产与呼吸消耗对策的异同;而其种内差异则反映了个体间生长状况的差异。本文的研究发现边材宽度在树种间和树种内均存在着显著的差异。7个树种的边材宽度波动在0.46~2.5 cm之间,其中被测针叶树边材宽度(1.96~2.55 cm)大于阔叶树(0.46~1.27 cm)。这与巨桉(Eucalyptus grandis)(0.9~4.8 cm)(Wilkins,1991)、蓝桉(E. globulus)(1.59~4.45 cm)(Miranda et al., 2006)、尾叶桉(E. urophylla)(1~3 cm)(周国逸等,2002)的边材宽度接近,却明显低于海岸松(Pinus pinaster)(5~20 cm)(Pinto et al., 2004;Knapic et al., 2005)、日本柳杉(>4 cm)(Yang et al., 1994)、黑云杉(Picea mariana)(>4 cm)(Hazenberg et al., 1991)。Sellin (1994;1996)发现,欧洲云杉(Picea abies)边材宽度与树冠等级有关,优势木可达7~9 cm,而被压木甚至低于2 cm。
3.2 心材和边材沿树干高度的变化所有树种的心材年轮数沿树干高度增加而递减,但是边材年轮数沿树高的变化却表现出不同的模式(图 2)。这与以往的研究结果一致,如:红松与北美短叶松(Larix laricina)(Yang et al., 1985)、辐射松(Pinus radiata)(Wilkes,1991)、海岸松(Pinto et al., 2004)和欧洲云杉(Longuetaud et al., 2006)一致,其边材年轮数均随高度增加递减;而兴安落叶松则与美洲落叶松(Pinus banksiana)(Yang et al., 1985)相同,其边材年轮数沿树干升高先增后减。阔叶树边材年轮数的垂直变化在以往研究中报道甚少。本研究中阔叶树的边材年轮数的垂直变化相对较小(图 2)。
本研究中所有树种的心材半径基本上随树干高度增大而减小,与Climent等(2003)报道的不规则模式(irregular pattern)不符(Wilkes,1991;Stokes et al., 2000;Berthier et al., 2001)。Climent等(2003)用相对树冠高度(枝下高占总树高的百分比)差异来解释加拿列松(Pinus canariensis)心材随高度的变化模式,认为树木摇动产生更多的乙烯可促进心材的形成,从而在摇动最剧烈的高度形成了较多的心材。而Pinto等(2004)和Knapic等(2005)却无法用相对树冠高度解释海岸松的心材变化格局。进一步分析表明,产生这2种不同结论的原因可能是取样方法不同,前者在优势木和亚优势木中随机取样;而后者却在林分中随机取样,包含了被压木。本研究按树木优势度取样,涵盖了优势木、中等木和被压木,而且风的影响在本地区并不显著。
树种间的心材半径虽然有一定变异性,但总体上沿树干随高度增大而减小,与树干外形基本一致(图 3)。但是,边材宽度随高度的变化比心材小得多,几乎所有树木的边材宽度在树干基部异常大,大约从1~2 m开始沿树高的变化不大(图 3)。这与以往的大部分研究结果一致(Climent et al., 2003;Knapic et al., 2005;Longuetaud et al., 2006)。虽然边材宽度从大约胸径开始一直到树冠变化很小,但由于树干的尖削度使边材横断面积随树高逐渐减小。
总之,树木种内和种间的心材和边材的生长特征差异,可以反映树木的生长状况、水分利用对策、物质生产和利用策略等生理生态特性的异同,这是探索树木水分关系和碳代谢机制、构建树木生长收获模型、木材工艺利用的基础。
汪矛, 张志农. 1998. 边材与心材. 生物学通报, 33(9): 13-14. |
周国逸, 黄志宏, Jim M, 等. 2002. 桉树人工林树液流动密度随边材径向深度的变化. 植物学报, 44(12): 1418-1424. DOI:10.3321/j.issn:1672-9072.2002.12.005 |
Bektas I, Alma M H, Yuksel A, et al. 2003. Influence of site on sapwood and heartwood ratios of Turkish Calabrian pine. Forest Products Journal, 53: 48-50. |
Berthier S, Kokutse A D, Stokes A, et al. 2001. Irregular heartwood formation in maritime pine (Pinus pinaster Ait): consequences for biomechanical and hydraulic tree functioning. Annals of Botany, 87: 19-25. DOI:10.1006/anbo.2000.1290 |
Bjorklund L. 1999. Identifying heartwood-rich stands or stems of Pinus sylvestris by using inventory data. Silva Fennica, 33: 119-129. |
Climent J, Chambel M R, Gil L, et al. 2003. Vertical heartwood variation pat terns and prediction of heartwood volume in Pinus canariensis SM. Forest Ecology and Management, 174: 203-211. DOI:10.1016/S0378-1127(02)00023-3 |
Climent J, Chambel M R, Perez E, et al. 2002. Relationship between heartwood radius and early radial growth, tree age, and climate in Pinus canariensis. Canadian Journal of Forest Research, 32: 103-111. DOI:10.1139/x01-178 |
Climent J, Gil L, Pardos J. 1993. Heartwood and sapwood development and its relationship to growth and environment in Pinus-canariensis Chr sm ex dc. For est Ecology and Management, 59: 165-174. DOI:10.1016/0378-1127(93)90077-Z |
Ericsson T, Fries A. 1999. High heritability for heartwood in north Swedish scots pine. Theoretical and Applied Genetics, 98: 732-735. DOI:10.1007/s001220051128 |
Gjerdrum P. 2003. Heartwood in relation to age and growth rate in Pinus sylvestris L. in Scandinavia. Forestry, 76: 413-424. DOI:10.1093/forestry/76.4.413 |
Hazenberg G, Yang K C. 1991. The relationship of tree age with sapwood and heart wood width in black spruce, Picea mariana (Mill) BSP. Holzforschung, 45: 317-320. DOI:10.1515/hfsg.1991.45.5.317 |
Knapic S, Pereira H. 2005. Within-tree variation of heartwood and ring width in maritime pine (Pinus pinaster Ait.). Forest Ecology and Management, 210: 81-89. DOI:10.1016/j.foreco.2005.02.017 |
Knapic S, Tavares F, Pereira H. 2006. Heartwood and sapwood variation in Acacia melanoxylon R. Br. trees in Portugal. Forestry, 79: 371-380. DOI:10.1093/forestry/cpl010 |
Longuetaud F, Mothe F, Leban J M, et al. 2006. Picea abies sapwood width: Variations within and between trees. Scandinavian Journal of Forest Research, 21: 41-53. DOI:10.1080/02827580500518632 |
Makela A, Vanninen P. 2001. Vertical structure of Scots pine crowns in different age and size classes. Trees-Structure and Function, 15: 385-392. DOI:10.1007/s004680100118 |
Miranda I, Gominho J, Lourenco A, et al. 2006. The influence of irrigation and fertilization on heartwood and sapwood contents in 18-year-old Eucalyptus globulus trees. Canadian Journal of Forest Research, 36: 2675-2683. DOI:10.1139/x06-130 |
Morling T, Valinger E. 1999. Effects of fertilization and thinning on heartwood area, sapwood area and growth in Scots pine. Scandinavian Journal of Forest Research, 14: 462-469. DOI:10.1080/02827589950154168 |
Pinto I, Pereira H, Usenius A. 2004. Heartwood and sapwood development within maritime pine (Pinus pinaster Ait.) stems. Trees-Structure and Function, 18: 284-294. DOI:10.1007/s00468-003-0305-8 |
Sellin A. 1994. Sapwood heartwood proportion related to tree diameter, age, and growth-rate in Picea abies. Canadian Journal of Forest Research, 24: 1022-1028. DOI:10.1139/x94-133 |
Sellin A. 1996. Sapwood amount in Picea abies L. Karst determined by tree age and radial growth rate. Holzforschung, 50: 291-296. DOI:10.1515/hfsg.1996.50.4.291 |
Shinozaki K, Yoda K, Hozumi K, et al. 1964. A quantitative analysis of plant form—the pipe model theory Ⅰ. Basic analyses. Japanese Journal of Ecology, 14: 95-105. |
Stokes A, Berthier S. 2000. Irregular heartwood formation in Pinus pinaster Ait. is related to eccentric, radial, stem growth. Forest Ecology and Management, 135: 115-121. DOI:10.1016/S0378-1127(00)00303-0 |
Wang C K. 2006. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. Forest Ecology and Management, 222: 9-16. DOI:10.1016/j.foreco.2005.10.074 |
Wang C, E W, Yang C, et al. 2007. Effects of provenance on development of sapwood and heartwood of Larix gmelinii trees. Forest Ecology and Management (in submission).
|
Wilkes J. 1991. Heartwood development and its relationship to growth in Pinus radiata. Wood Science and Technology, 25: 85-90. |
Wilkins A P. 1991. Sapwood, heartwood and bark thickness of silviculturally treated Eucalyptus grandis. Wood Science and Technology, 25: 415-423. DOI:10.1021/es00015a006 |
Woeste K E. 2002. Heartwood production in a 35-year-old black walnut progeny test. Canadian Journal of Forest Research, 32: 177-181. DOI:10.1139/x01-177 |
Yang K C, Chen Y S, Chiu C, et al. 1994. Formation and vertical-distribution of sapwood and heartwood in Cryptomeria japonica D Don. Trees-Structure and Function, 9: 35-40. |
Yang K C, Hazenberg G, Bradfield G E, et al. 1985. Vertical variation of sap wood thickness in Pinus banksiana Lamb. and Larix laricina (Duroi) Koch, K. Canadian Journal of Forest Research, 15: 822-828. DOI:10.1139/x85-133 |