 2008, Vol. 44
2008, Vol. 44文章信息
- 田呈明, 赵鹏, 曹支敏.
- Tian Chengming, Zhao Peng, Cao Zhimin.
- 细胞壁降解酶在落叶松-杨栅锈菌与寄主互作过程中的作用
- Role of Cell Wall-Degrading Enzymes in Interaction of Poplar and Melampsora larici-populina
- 林业科学, 2008, 44(5): 79-83.
- Scientia Silvae Sinicae, 2008, 44(5): 79-83.
-
文章历史
- 收稿日期:2006-11-15
-
作者相关文章


2. 西北农林科技大学林学院 杨凌 712100
2. College of Forestry, Northwest A & F University Yangling 712100
杨树叶锈病(Melampsora larici-populina)是杨树苗期和幼树阶段的重要病害之一,病原菌变异快、寄主种类多、损失严重,制约杨树商品林和生态林的发展(田呈明等,1999)。目前,许多学者对该病害的致病性分化和寄主抗性等方面进行了研究,但缺乏有关病菌致病机理的研究(Pei et al., 2005),而病原菌致病性机理研究则为抗性育种提供重要的理论依据。
植物细胞壁是病原菌入侵、扩展的最大屏障。因此,在入侵过程中病菌产生的一些细胞壁降解酶是决定病原菌成功入侵植物的重要因子,这些相关细胞壁降解酶与病原菌的致病性有一定的关系(赵蕾等,2002)。许多研究表明,非专性寄生菌在入侵过程中产生大量的细胞壁降解酶,而有关专性寄生菌细胞壁降解酶的研究却很少(李宝聚等,2000;芦晓飞,2005)。对小麦锈菌、菜豆锈菌等研究后认为,在专性寄生菌入侵寄主的过程中的确存在细胞壁降解酶,并对入侵发挥重要的作用(芦晓飞,2005;Xu, et al., 1997)。芦晓飞(2005)对小麦与条锈菌互作系统中细胞壁降解酶的研究发现,聚甲基半乳糖醛酸酶(PMG)、果胶甲基反式消除酶(PMTE)、羧甲基纤维素酶(CX)、β-葡萄糖苷酶(β-G)等四种降解酶在小麦条锈菌侵染过程中发挥了重要的作用,但在锈菌等寄生菌与寄主互作系统中上述细胞壁降解酶的含量较低,并不像非专性寄生菌那样能在侵染过程中产生大量的细胞壁降解酶。前人对栅锈菌与寄主互作过程中寄主酶的变化进行了研究(魏益宁,1984;樊自红,1989),而有关互作过程中病原菌是否产生细胞壁降解酶及其作用仍缺乏研究。因此,通过研究栅锈菌与不同抗性杨树品种互作系统中的细胞壁降解酶,揭示细胞壁降解酶在落叶松—杨栅锈菌侵染过程中所起的作用,为科学认识栅锈菌致病机理奠定理论基础。
1 材料和方法 1.1 供试菌种和树种供试杨树无性系为川杨(Populus szechuanica)和太白杨(P. purdomii)。采集1年生的插条,盆栽并放置于温室内培养。试验用菌种Bq(采自北京延庆)和Sb(采自四川宝兴)为我国落叶松—杨栅锈菌的生理小种的单孢菌系,由西北农林科技大学林学院森林病理实验室提供。两个单孢菌系中Bq在太白杨上反应型为(4),在川杨上则为(0);Sb在太白杨上反应型为(4),在川杨上为(1)。菌种的繁殖、保存、反应型测定标准和方法参照曹支敏等(1998)的方法。
1.2 接种和取样方法选取叶龄指数(leaf plastochron index,LPI)为5~8的叶片作为接种叶片,LPI值测定方法参照Larson等(1971)的方法,保存的菌种经过活化后配成浓度为2×104个·mL-1的孢子悬浮液,用玻璃棒沾取夏孢子悬浮液均匀地涂抹于叶背面,接种密度为100 mL·m-2,苗子接种后保湿24 h,然后取出放置在温室中,以未接种的健康苗子为对照。在接种前取健康叶片并测定其细胞壁降解酶活性,在接种后的1、2、3、5、7 d(days post-inoculation, dpi)分别各取样一次,测定细胞壁降解酶的变化。不接种的健康叶片以相同的方法取样作为对照。
1.3 细胞壁降解酶测定细胞壁降解酶提取方法参照芦晓飞(2005)的方法,应用分光光度计法;参照芦晓飞(2005)、李宝聚等(2000)的方法,分别测定聚甲基半乳糖醛酸酶(PMG)、果胶甲基反式消除酶(PMTE)、羧甲基纤维素酶(CX)、β-葡萄糖苷酶(β-G),取样量为1 g, 所有处理均重复3次。
2 结果和分析 2.1 聚甲基半乳糖醛酸酶(PMG)的活性变化在接种24 h后开始,川杨和太白杨在接种不同致病性的菌样之后体内PMG活性均有所增加,接种处理比未接种处理的PMG活性变化明显,但两种寄主上的接种处理的PMG与对照的差值即ΔPMG具体变化各不相同。在接种Bq菌株的川杨体内,接种后第2天ΔPMG开始活性变化明显,并在接种后第2天和接种后第5天存在2个活性高峰,其中接种后第5天达到最大活性高峰,接种处理比未接种处理要大0.440 3 U·mg-1;接种Sb菌株之后的第1天,ΔPMG活性就开始增加,在接种后第2天时接种处理要比未接种处理大0.176 8 U·mg-1,达到活性高峰。在太白杨上,接种Bq后的第1天开始ΔPMG活性明显发生变化,在接种后第5天接种处理比未接种的要大1.522 0 U·mg-1, 达到最大活性高峰;接种Sb的第3天时接种处理比对照要大0.402 7 U·mg-1,达到活性最大峰值。在感病品种太白杨上ΔPMG活性值较抗病品种川杨上的大2.3~3.5倍。不仅如此两种杨树在接种Bq后ΔPMG活性要比接种Sb的大1.2~4.4倍(图 1、2)。

|
图 1 受落叶松-杨栅锈菌入侵后川杨叶片内PMG活性变化 Figure 1 Changes of PMG activity in Populus szechuanica infected by Melampsora larici-populina 接种时间 Inoculation time/d □CK ■Bq 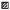 Sb Sb |

|
图 2 受落叶松-杨栅锈菌入侵后太白杨叶片内PMG活性变化 Figure 2 Changes of PMG activity in P.purdomii infected by Melampsora larici-populina 接种时间 Inoculation time/d □CK ■Bq Sb |
抗病品种和感病品种中PMTE活性变化差异很大,而且不同菌种与寄主组合内的PMTE与对照的PMTE差值即ΔPMTE的变化趋势也各不相同。在川杨上接种Sb菌样后的第1天,ΔPTME活性就开始增加,在接种后第3天时接种处理比未接种处理的ΔPMTE活性要大0.280 9 U·mg-1,达到活性峰值。之后,ΔPMTE活性开始逐渐下降;接种Bq后第1天ΔPMTE活性比对照的要大0.181 9 U·mg-1,达到活性高峰,在接种后第3天时又出现另一个峰值0.141 0 U·mg-1,之后逐渐下降。在感病品种太白杨上,接种Sb后ΔPMTE活性逐渐开始上升一直到接种后第7天,其中并没有出现活性最大峰值;而接种Bq后ΔPMTE活性逐渐增加,到接种后第3天时接种处理比对照要大0.747 3 U·mg-1,达到活性峰值。之后又逐渐开始降低,整个过程中出现一个活性高峰。从活性峰值可以看出在感病品种太白杨上ΔPMTE的活性比在抗病品种川杨要大2.7~3.2倍(图 3、4)。

|
图 3 受落叶松-杨栅锈菌入侵后川杨叶片内PMTE活性变化 Figure 3 Changes of PMTE activity in P. szechuanica infected by M. larici-populina 接种时间 Inoculation time/d □CK ■Bq Sb |

|
图 4 受落叶松-杨栅锈菌入侵后太白杨叶片内PMTE活性变化 Figure 4 Changes of PMTE activity in P. purdomii infected by M. larici-populina 接种时间 Inoculation time/d □CK ■Bq Sb |
接种后CX活性的变化也存在这明显的差异,尤其是其接种处理与未接种处理的差值ΔCX在川杨和太白杨上变化差异很大。在川杨上,接种两种菌样后ΔCX活性变化趋势基本上一致,在接种后第2天时都达到活性最高峰,之后就逐渐降低。接种Sb后活性高峰出现时接种处理比对照大0.089 5 U·mg-1,而接种Bq后接种处理比对照大0.056 4 U·mg-1;在感病品种太白杨上接种两种菌样后变化趋势基本上趋于一致,先增大到峰值之后就逐渐降低,但是峰值出现的时间有早有晚。接种Sb后ΔCX在第3天时出现峰值,接种处理比对照大0.286 29 U·mg-1。而接种Bq后ΔCX第2天时早就出现峰值,接种处理比对照要大0.278 5 U·mg-1。总的来说,CX活性在抗病品种中较感病品种中的活性低(图 5、6)。

|
图 5 受落叶松-杨栅锈菌入侵后川杨叶片内CX活性变化 Figure 5 Changes of CX activity in P. szechuanica infected by M. larici-populina 接种时间 Inoculation time/d □CK ■Bq Sb |

|
图 6 受落叶松-杨栅锈菌入侵后太白杨叶片内CX活性变化 Figure 6 Changes of CX activity in P. purdomii infected by M. larici-populina 接种时间 Inoculation time/d □CK ■Bq Sb |
川杨在接种Sb后的第一天开始β-G活性变化就比较明显,接种处理比对照要大0.045 1 U·mg-1, 在接种后第2天Δβ-G活性达到峰值,接种处理比对照要大0.062 3 U·mg-1,之后活性逐渐降低。在接种Bq后Δβ-G第3天时活性才到达峰值,接种处理比对照大0.033 3 U·mg-1,之后同样有降低的趋势。在太白杨上,接种Sb、Bq后Δβ-G活性变化一致,在接种后第一天开始活性就逐渐增加,均在接种后第3天时出现活性峰值,之后活性又逐渐开始降低。出现峰值时接种Sb的太白杨中β-G活性比对照大0.179 1 U·mg-1,而接种Bq的太白杨中β-G活性比对照大0.210 5 U·mg-1。

|
图 7 受落叶松-杨栅锈菌入侵后川杨叶片内β-G活性变化 Figure 7 Changes of β-G activity in P. szechuanica infected by M. larici-populina 接种时间 Inoculation time/d □CK ■Bq Sb |

|
图 8 受落叶松-杨栅锈菌入侵后太白杨叶片内β-G活性变化 Figure 8 Changes of β-G activity in P. purdomii infected by M. larici-populina 接种时间 Inoculation time/d □CK ■Bq Sb |
1) 对Melampsora larici-populina与寄主杨树互作过程中相关细胞壁降解酶的动态变化研究发现,在受落叶松—杨栅锈菌侵染之后,无论抗病品种还是感病品种中均测出有聚甲基半乳糖醛酸酶(PMG)、果胶甲基反式消除酶(PMTE)、羧甲基纤维素酶(CX)、β-葡萄糖苷酶(β-G)的存在,但是活性较非专性寄生菌如黄瓜黑星病菌(Cladosporium cucumerinum)、番茄霜霉病菌(Botrytis cinerea)、玉米弯孢霉叶斑病菌(Curvularia lunata)等入侵寄主时产生的降解酶活性要低(陈夕军等,2006;冯晶等,2002;李宝聚等,2000;2003)。同时,几种酶活性在感病品种太白杨和抗病品种川杨中也有差异。在感病品种中这些细胞壁降解酶的活性比抗病品种中的高,其活性峰值是抗病品种中的2~5倍。这说明栅锈菌同小麦锈菌等其他专性寄生菌一样,在入侵寄主过程中产生这几种细胞壁降解酶,且细胞壁降解酶在致病性分化中发挥着一定的作用。
2) 植物病原菌在入侵寄主的过程中为了穿透并进入植物组织,必须分泌一系列的细胞壁降解酶来消解植物机械屏障从而达到摄取营养物质的目的。在病原菌分泌的这些降解酶中PMG、PMTE均是以果胶质为基质的水解酶,而CX、β-G能分解纤维素并使其最终水解成葡萄糖(赵蕾等,2002),几种降解酶的综合作用促进病原菌对营养的吸收。与非寄生及半寄生菌依靠破坏寄主细胞来摄取营养的方式不同,专性寄生菌如锈菌、白粉菌在入侵过程中只在侵染点附近发生细胞降解。可能是因为病原菌入侵中严格控制着降解酶的表达,达到成功入侵、定殖而不破坏寄主细胞的目的,从而维持着组织的完整性(Christoph et al., 1999)。田呈明等(2002)在对栅锈菌的入侵过程中的超微结构研究中发现,栅锈菌的侵入并不对寄主细胞产生大的损伤,入侵菌丝紧贴细胞壁在细胞间隙中扩展。本研究发现,接种栅锈菌后,抗病和感病品种中PMG和PMTE的活性高,而CX和β-G的活性则相对较小。这可能是因为PMG、PMTE主要降解寄主细胞壁胞间薄层和初生壁中的果胶物质,促使入侵菌丝能快速在细胞壁间隙中扩展但不破坏寄主细胞壁,而CX和β-G降解细胞壁次生壁中的纤维素,在栅锈菌扩展、定殖中也发挥一定的作用。此外,PMG、PMTE同CX、β-G的活性差异一定程度上说明果胶酶在栅锈菌入侵过程中的作用较纤维素酶显著。
3) Deising等(1995)和Heiler等(1993)对Uromyces viciae-fabae的研究发现,双核夏孢子入侵寄主过程中分泌一系列细胞壁降解酶时具有高度的时序性。纤维素酶在附着胞形成过程中开始分泌,而PMTE则在侵染菌丝扩展过程中分泌,聚半乳糖醛酸裂合酶分泌时期则在吸器母细胞分化过程之中。无论抗病组合还是感病组合,几种细胞壁降解酶所表达的时空阶段基本上是一致的。本研究发现,同样在栅锈菌和杨树互作系统中,无论在川杨和太白杨中,ΔPMG均在接种后第1天和第5天时出现两个活性高峰,ΔPMTE在接种后第3天时出现活性高峰(太白杨上接种Bq时高峰出现在接种后第1天),ΔCX在接种后第2天时出现活性高峰(太白杨上接种Sb时活性高峰出现在接种后第3天),Δβ-G在接种后第3天时出现活性高峰(川杨接种Sb时活性高峰出现在接种后第2天),这说明栅锈菌细胞壁降解酶分泌同样也具有高度的时序性。栅锈菌在入侵过程中严格控制这几种细胞壁降解酶的表达,而且时空表达并不受寄主防御体系的影响。无论抗病组合还是感病组合,几种细胞壁降解酶所表达的时空阶段基本上是一致的。
4) 植物细胞壁降解酶不仅是病原菌主要致病因子,而且也是寄主抗病反应的主要诱导因子(赵蕾等,2002)。在非专性病原菌黄瓜黑星病菌(Cladosporium cucumerinum)与寄主互作后的细胞壁降解酶研究中认为,抗病品种在感病前期出现的相关细胞壁降解酶酶活高峰,可能诱发寄主防卫反应的发生,这些降解酶起激发子的作用(李宝聚等,2000)。在本研究中,抗病组合之中PMTE、PMG、CX在接种后的1~2天也出现一个小的活性高峰,可能是这些降解酶与非专化病原菌产生的细胞壁降解酶作用相类似,即作为寄主抗病反应的诱导因子诱发寄主防卫反应,从而产生一系列保护酶抑制病原菌的侵染。
5) 细胞壁降解酶与落叶松—杨栅锈菌的侵染、扩展有一定的联系。但是在不同温度下致病菌产生的致病酶活性不同,病原菌产生的细胞壁降解酶的活性随着温度的升高会降低,而随着湿度的增高酶活却增加(李宝聚等,2003)。因此在整个研究过程中,由于温度、湿度的变化对这几种细胞壁降解酶的作用和表达时间会有一定的影响,在本试验中,随着接种时间的延续,接种处理和对照中细胞壁降解酶出现的差值能定性的分析出细胞壁降解酶在专性寄生菌入侵寄主过程中作用,但是却很难定量分析其在具体的时间段中所产生的影响。因此确定合理的温度和相对湿度能揭示细胞壁降解酶在栅锈菌与杨树互作过程中的时空表达。此外,细胞壁降解酶对寄主超微结构的影响及作用位点的确定以及果胶酶、纤维素酶在侵染中的协同作用尚须进一步的研究。
曹支敏, 李振岐, 胡锦江. 1998. 落叶松-杨栅锈菌(Melampsora larici-populina Kleb.)生理分化研究. 西北林学院学报, 13(1): 53-57. |
陈夕军, 张红, 徐敬友, 等. 2006. 水稻纹枯病菌胞壁降解酶的产生及致病作用. 江苏农业学报, 22(1): 24-28. DOI:10.3969/j.issn.1000-4440.2006.01.006 |
樊自红. 1989. 过氧化物酶和苯丙氨酸解氨酶与毛白杨抗锈性的关系. 植物病理学报, 19(2): 95-100. DOI:10.3321/j.issn:0412-0914.1989.02.003 |
冯晶, 高增贵, 薛春生, 等. 2002. 玉米弯孢霉叶斑病菌产生的细胞壁降解酶的致病作用研究. 杂粮作物, 22(3): 164-166. DOI:10.3969/j.issn.2095-0896.2002.03.015 |
李宝聚, 周长力, 赵奎华, 等. 2000. 黄瓜黑星病菌致病机理的研究Ⅱ细胞壁降解酶及其在致病中的作用. 植物病理学报, 30(1): 13-18. DOI:10.3321/j.issn:0412-0914.2000.01.003 |
李宝聚, 陈立芹. 2003. 温湿度调控对番茄灰霉病菌产生的细胞壁降解酶的影响. 植物病理学报, 33(3): 209-212. DOI:10.3321/j.issn:0412-0914.2003.03.004 |
芦晓飞. 2005.小麦—条锈菌互作系统中几种蛋白因子的研究.西北农林科技大学硕士学位论文. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y752867
|
田呈明, 李振岐, 康振生. 1999. 青杨叶锈病研究进展. 西北林学院学报, 14(2): 81-88. |
田呈明, 梁英梅, 康振生, 等. 2002. 青杨叶锈病菌(Melampsora larici-poulina Kleb.)侵染过程的超微结构研究. 植物病理学报, 32(1): 71-78. DOI:10.3321/j.issn:0412-0914.2002.01.013 |
魏益宁. 1984. 毛白杨叶片受马格栅锈菌侵染以后多酚氧化酶和过氧化物酶活性及同功酶谱带变化趋势的研究. 北京林业大学学报, 17(3): 73-92. |
赵蕾, 张天宇. 2002. 植物病原菌产生的降解酶及其作用. 微生物学通讯, 29(1): 89-93. |
Christoph S S, Gerhard A W. 1999. Cellulase in the host-parasite system Phaseolus vulgaris L.-Uromyces appendiculatus (Pers.) Schroet. European Journal of Plant Pathology, 105: 285-295. DOI:10.1023/A:1008763929057 |
Deising H, Frittrang A K, Kunz S, et al. 1995. Regulation of pectin methyl esterase and polygalacturonate lyase activity during differentiation of infection structures in Uromyces viciae-fabae. Microbiology, 141: 561-571. DOI:10.1099/13500872-141-3-561 |
Heiler S, Mendgen K, Deising H. 1993. Cellulotic enzymes of the obligately biotrophic rust fungus Uromyces viciae-fabae are regulated differentiation-specifically. Mycol Res, 97: 77-85. DOI:10.1016/S0953-7562(09)81116-7 |
Larson P R, Isebrands J G. 1971. The plastochron index as applied to development al studies of cottonwood. Can J Forest Res, 1: 1-11. DOI:10.1139/x71-001 |
Pei M H, McCracken A R. 2005. Rust diseases of willow and poplar. CABI Publishing, 1-264.
|
Xu H, Mendgen K. 1997. Targeted cell wall degradation at the penetration site of cowpea rust basidiosporelings. The American Phytopathological Society, 10(1): 87-94. |