 2008, Vol. 44
2008, Vol. 44文章信息
- 孙晓梅, 张守攻, 王笑山, 齐力旺, 王万义, 陈宝丰.
- Sun Xiaomei, Zhang Shougong, Wang Xiaoshan, Qi Liwang, Wang Wanyi, Chen Baofeng.
- 日本落叶松×长白落叶松杂种组合间生根性状及幼林生长的遗传变异
- Genetic Variation on Rooting Ability of Stem Cutting and Juvenile Growth of Larix kaempferi × L. olgensis Interspecific Hybrids
- 林业科学, 2008, 44(4): 41-47.
- Scientia Silvae Sinicae, 2008, 44(4): 41-47.
-
文章历史
- 收稿日期:2007-10-26
-
作者相关文章



2. 辽宁省清源县大孤家国营林场 清源 113305
2. Dagujia Forest Farm in Qingyuan County Qingyuan 113305
杂种优势的发现和杂种在速生、抗逆方面的出色表现,使得杂种利用成为林木遗传改良工作的重要组成部分。在林木中落叶松种间杂交育种的研究工作开展得比较早。自1990年欧洲落叶松(Larix decidua)和日本落叶松(L. kaempferi)的天然杂种在欧洲被发现以来,很多国家致力于日本落叶松与当地落叶松种的杂交育种,并推出了杂种优势明显、对生产有影响的欧(欧洲落叶松)×日(日本落叶松)、日×欧、日×兴(兴安落叶松L. gmelinii)等种间杂种(Macdonald et al., 1957; Baltunis et al., 1998; Li et al., 1994;Matyssek et al., 1987;Luc, 2004;王战等,1992)。我国从20世纪70年代开始相继开展了日×长(长白落叶松L. olgensis)、长×日、日×华(华北落叶松L. principis-rupprechtii)、日×兴、兴×日、兴×长等的杂交育种工作,通过遗传测定,普遍认为在东北地区日×长的多数杂交组合由于集合了双亲的优良性状,在生长、干形、材性和抗逆(抗寒、抗病、抗鼠害)等方面表现出明显的杂种优势(潘本立等,1981;杨书文等,1985;王景章等,1980;张含国等,1998;2005;孙晓梅等,2008)。然而由于种间花期不遇、结实周期长及大小年现象明显等原因,营建杂种或双系种子园无法满足商业化生产对杂种种子的需求。无性繁殖技术的发展,为杂种利用开辟了一条高效新途径,并形成了以人工控制授粉有性配制杂种为基础、采穗圃经营为关键、扦插繁殖利用为手段的落叶松杂种规模利用配套技术体系(Kyle et al., 1997;王笑山等,2000)。目前,相关的研究主要集中在生根技术方面(孙晓梅等,2006;Kyle et al., 1997;John,1979;Mason,1989),而对落叶松杂种组合间生根变异方面的研究以及生根与生长性状的联合选择却少有报道。本文利用在辽宁大孤家杂种实生采穗园内不同母本与同一多父本混合花粉及同一母本与不同单一父本花粉2种授粉方式获得的12个日×长种间杂交组合及相应母本自由授粉家系为试材,开展杂种子代生根、幼林生长性状遗传变异研究,揭示不同组合间生根及生长性状的父、母本效应,并利用隶属函数法选出生根最优、生长最优及生根与生长兼优的杂交组合。
1 材料与方法 1.1 试验材料试验用日×长杂种组合及其母本自由授粉家系半木质化插穗采自辽宁省清源县大孤家林场落叶松杂种采穗园。4个日×长多父本混合授粉(4个长白落叶松无性系花粉等量混合,花粉采自辽宁哈达种子园)杂种(日永2×长混、日永8×长混、日清4×长混、日永85×长混,其中日永2、日永8、日清4、日永85为辽宁大孤家日本落叶松种子园无性系编号)及其4个对应母本自由授粉家系为1995年杂交采种,1996年播种育苗,1997年建采穗园;同母本不同父本日×长杂种(日永85×长10、日永85×长11、日永85×长12、日永85×长13、日永85×长14、日永85×长27、日永85×长3、日永85×长4C,其中长11、长10、长12、长13、长14、长27、长3和长4C为长白落叶松无性系编号,花粉采自辽宁哈达种子园)及日永85自由授粉家系为1996年杂交采种,1997年春播种育苗,1998年建采穗园。采穗园定植株行距1 m×1 m,母株采用高柱式整形修剪。
1.2 试验设计与方法扦插生根试验,包括4个日×长多父本混合授粉杂交组合、8个同母本不同父本日×长杂交组合及4个相对应的同母本日本落叶松自由授粉家系,2002年6月下旬分组合(家系)各从10~30株以上母株上采穗扦插,穗长15 cm。扦插试验采用随机区组设计,4次重复,每小区插50条插穗。相同组合(家系)插穗充分混合后,经清水浸基部30 min后扦插,深3 cm,密度666穗·m-2 (5 cm×3 cm),基质为纯净河砂。采用全光雾插技术进行插床管理(王笑山等,1995)。当年秋季分区调查每穗是否生根及偏根、生根量、最长根长,计算小区插穗生根率和偏根率。2003年春生根插穗换床继续培育1年。
2004年春用2年生扦插苗造林。试验林采用完全随机区组设计,4次重复,16株方形小区。穴状整地,整地规格60 cm×60 cm×40 cm,株行距2 m×2 m,抚育方式采用3年5次(2:2:1)。在每年生长停止后定株调查树高、地径,并于造林当年和第3年统计各组合(家系)的造林成活率和保存率。
1.3 统计分析方法各性状方差分析、家系遗传力、性状间遗传相关和表型相关系数计算公式见相关文献(Svensson et al., 1999;Falconer, 2000)。方差分析采用SAS/STAT 6.12软件中PROC ANOVA或GLM过程计算,各方差分量用PROC VARCOMP过程中TYPE1方法计算(高惠璇等,1997),其中生根率、偏根率及成活率经反正弦变换。利用双因素分析和LSD多重比较方法研究不同组合间生根和生长性状的差异,利用Dunnett法进行杂交组合与日本落叶松自由授粉家系间差异显著性比较,并利用隶属函数法(陈荣敏等,2002)选出最优生根、生长组合(家系)。
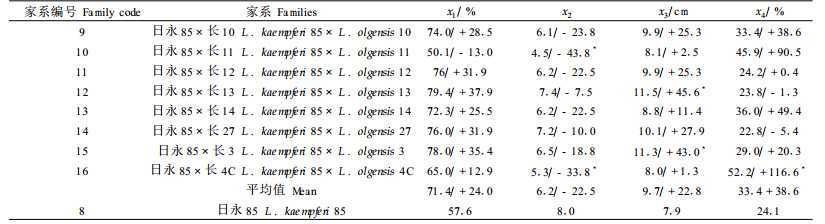
2 结果与分析 2.1 日×长杂交组合生根性状的遗传变异 2.1.1 多父本混合授粉杂种子代与母本自由授粉子代生根性状的变异及母本效应比较多父本混合授粉杂种子代与母本自由授粉子代生根性状(表 1)发现,杂种组合的生根率和偏根率普遍高于对应的母本自由授粉家系(日永2除外,日永2×长混的各生根性状均低于母本),平均生根量大多低于对应的母本自由授粉家系,而偏根率普遍高于对应母本自由授粉家系(日清4除外,日清4×长混的各生根性状均优于母本),说明杂交组合较母本对照的扦插生根率有所提高,而生根质量有所下降。方差分析结果(表 2)表明,母本自由授粉家系(纯种)与杂交组合(杂种)间只有平均每穗生根量存在显著差异,其他性状差异不显著,另据各组合与同母本对照成对比较结果,日永2×长混的生根率和生根量显著低于日永2,日永85×长混的最长根长显著高于日永85。而4个不同母本杂交组合间生根性状没有显著差异,说明组合中的母本效应并不明显。所有参试家系(包括纯种和杂种)间各生根性状均存在显著差异,家系遗传力在0.74~0.87之间,说明生根性状受中等以上强度的遗传控制,开展生根性状的选择能够取得良好的效果。从表型和遗传变异系数来看,偏根率的变异系数最大,达61.67%,最长根长的变异系数最小, 为16.93%。
|
|

|
|

由表 3可知,同母本不同父本日×长的大多数杂种组合的生根率和最长根长均高于母本自由授粉家系,平均超母本优势分别达24%和22.8%;而生根质量却不如自由授粉家系,生根量少(22.5%)和偏根率高(38.6%),说明杂种生根量受父本长白落叶松的影响,不如日本落叶松纯种,从而导致杂种的偏根率也偏高,这与长白落叶松较日本落叶松更难生根、生根量少有关。方差分析结果(表 4)表明,不同组合间生根量、最长根长差异达显著水平,生根率也达10%的显著水平,这种组合间的性状差异主要为父本效应引起的。其中日永85×长11和日永85×长4C的生根量显著低于母本对照,日永85×长13和日永85×长3的最长根长、日永85×长4C的偏根率显著高于日永85。通过估计性状的遗传参数发现,各生根性状的家系遗传力在0.68~0.81之间(表 4),说明生根性状受中等以上强度的遗传控制,因此开展不同杂种组合生根性状的选择能够取得良好的效果。从表型和遗传变异系数来看,同母本的日×长杂交组合的变异系数比混合授粉的变异要小,生根性状中也是偏根率的变异系数最大(46.20%),平均最长根长最小(18%)。
|
|
|
|

4个混合授粉杂交组合的生长量整体上优于母本自由授粉子代(日永85×长混和日永8×长混组合的生长量和保存率超过母本对照,日永2×长混与母本生长量相当,日清4×长混稍差于母本对照),5年生树高、地径和当年高分别提高7.0%、6.5%和3%,杂种优势并不十分明显,但保存率却比母本对照提高22.2%(表 5)。方差分析结果(表 6)表明,母本自由授粉家系(纯种)与杂交组合(杂种)间的3年生树高、地径及造林保存率存在显著差异,其他性状差异不显著,另据各组合与同母本对照的成对比较结果,日永8×长混3年生、4年生地径、日永85×长混造林成活率、5年生树高和地径均显著高于母本对照。4个不同母本杂交组合间地径和树高均存在显著或极显著变异,说明组合中生长性状的母本效应明显,开展杂种子代的生长性状选择具有很大的潜力。除了4年生当年高生长的家系遗传力较低以外,其他各年生长性状的家系遗传力均在0.78~0.92之间,说明这些性状受较强的遗传控制。
|
|

|
|
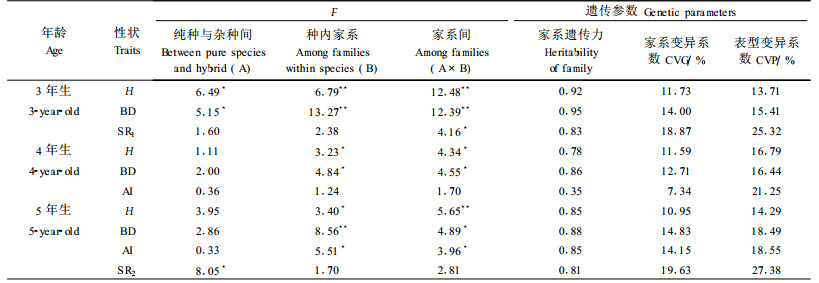
不同父本日永85×长杂种组合的3~5年生长量、造林成活率及保存率均超过母本对照,5年生树高、地径、当年高及保存率分别提高35.5%、28.6%、52.1%和59.4%。其中,日永85×长4C和日永85×长14组合的5年生树高和地径超过对照50%以上,显著高于对照(表 7)。方差分析结果表明,除了4年生地径和5年生当年高生长外,其他各年生长量、造林成活率及保存率组合间均达显著水平,这种变异主要是由父本效应引起的,由此说明开展杂交亲本及杂交组合间选择十分必要。除了4年生地径的家系遗传力(0.30)偏低以外,其他各年均在0.5以上(表 8),表明这些性状受中度以上的遗传控制,进行组合间选择能够取得较好的效果。从表型和遗传变异系数来看,造林成活率和3年保存率的变异系数最大,在32%以上,各年生长性状的变异幅度相对较小,在12%~29%之间(表 8)。
|
|

|
|
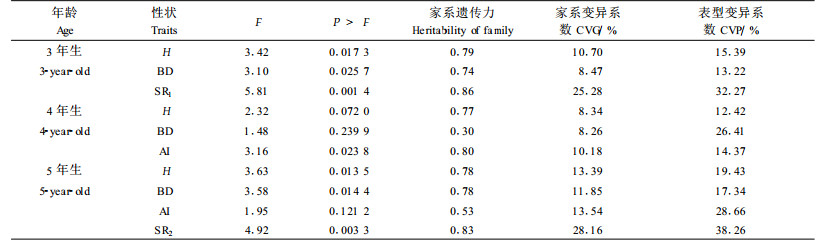
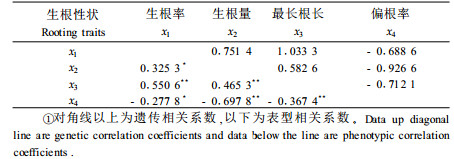
研究不同性状间的相关对了解这些性状的遗传控制,简化选择程序,做出选择决策有着重要意义(王笑山等,1995;马常耕等,1994)。各参试组合(家系)生根性状间存在着紧密的相关关系,且这些相关关系主要受遗传因素控制(表 9)。生根率、生根量和最长根长之间存在着显著正相关,而偏根率与这3个性状之间存在着显著负相关关系。其中,偏根率与生根量的相关最紧密,相关系数高达-0.70,即生根量越少,根系偏根率越高。这一结果与马常耕等(1994)、王笑山等(1995)对日本落叶松及Farmer等(1992)对美洲落叶松(Larix laricina)的研究结果一致。因此,为了节省调查时间、简化选择程序,在进行生根力选择时可将每穗生根量作为主导调查因子,生根率通过统计生根插穗数获得。由表 10可知,生根性状与各年幼林生长性状之间相关均不紧密,可以开展生根与生长性状的独立选择。
|
|
|
|

本研究选择组合(家系)间存在显著变异的生根率和生根量(偏根率与生根量紧密相关,最长根长与生根率紧密相关)作为生根性状的评价指标,选择5年生树高、地径和保存率作为生长性状的主要评价指标,采用隶属函数法(也称为多性状综合评价法)开展生根与生长性状的综合评价(表 11)。根据各组合(家系)计算的隶属函数值选出4个生根最优的组合(家系)(入选率为25%),依次为日永2、日永85×长13、日永85×长27、日永85×长3,入选家系的平均生根率和每穗生根量分别为81.4%和7.4条,分别比参试组合(家系)的平均值提高15.7%、20.7%;选出4个生长最优、生长和生根兼优的家系,依次为日永85×长14、日永8×长混、日永85×长4C和日永85×长13,入选家系5年生(含2年苗龄)平均树高、地径和保存率分别为2.32 m、3.39 cm和90.63%,分别比参试组合(家系)的平均值提高16.0%、19.8%和24.6%,比参试的4个日本落叶松对照家系的平均值提高24.7%、28.9%和55.6%,而生根性状与其基本一致,生根率还略有提高。
|
|
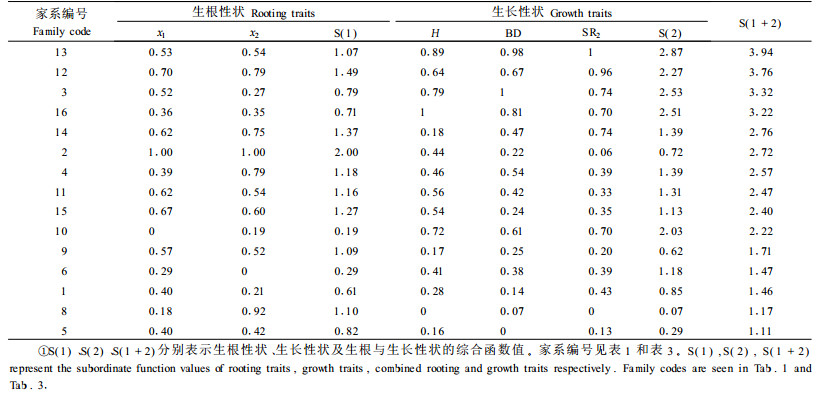
日×长杂种子代较母本自由授粉子代生根率有所提高,生根质量有所下降(生根量少、偏根率高),生长量整体上优于母本自由授粉子代。由此可以看出,日×长种间杂种优势十分明显,但也不是所有的杂交组合都必然优于母本,开展组合间的选择十分必要。由于当初没有采集对应的长白落叶松自由授粉家系的种子,本试验缺少父本对照,也就无法分析杂交后代对比父本的杂种优势,但长白落叶松的生长量远不如日本落叶松,生根力也不及日本落叶松,所以杂种超父本优势会更加明显。
无论是生根性状还是幼林生长性状,同母本与不同父本日×长杂交组合间存在着显著的遗传变异,即父本效应十分明显,为落叶松种间交配中亲本选择及更优杂交组合的选择提供了依据;不同母本与多父本混合授粉组合间只有生长性状存在着显著变异,体现了一定的母本效应,但生根性状的母本效应不明显,这可能是由于参试组合少,且是混合授粉。因此,有待进一步开展多个日本落叶松与长白落叶松亲本材料的双列杂交试验以获得更加丰富而有价值的遗传信息。主要生根和生长性状的家系遗传力在0.68~0.95之间,说明生根性状受中等以上强度的遗传控制,开展组合间生根和生长性状的选择能够取得良好的效果。
各参试组合(家系)生根性状间存在着紧密的相关关系,其中生根率、生根量和最长根长之间存在着显著正相关,而偏根率与这3个性状之间存在着显著负相关关系,为简化调查和选择程序可只调查每穗生根量这一主导生根因子。但生根性状与各年幼林生长性状之间相关不紧密,可以开展生根与生长性状的独立选择。
选出4个生根与生长兼优的家系(日永85×长14、日永85×长13、日永8×长混和日永85×长4C),5年生平均树高、地径和保存率分别比参试平均值提高16.0%、19.8%和24.6%,比4个母本对照提高24.7%,28.9%和55.6%,而生根性状与其基本一致。
陈荣敏, 杨学举, 梁凤山, 等. 2002. 利用隶属函数法综合评价冬小麦的抗旱性. 河北农业大学学报, 25(2): 7-9. |
Falconer D S. 2000. 数量遗传学导论. 储明星, 译. 北京: 中国农业科技出版社.
|
高惠璇, 耿直, 李贵斌, 等. 1997. SAS系统SAS/STAT软件使用手册. 北京: 中国统计出版社.
|
马常耕, 王笑山, 王建华, 等. 1994. 日本落叶松插穗生根能力的变异和选择效应. 林业科学, 30(2): 97-103. |
潘本立, 艾正明, 韩承伟, 等. 1981. 落叶松立木杂交方法及育种优势的研究. 林业科学, 17(3): 325-330. |
孙晓梅, 张守攻, 王笑山, 等. 2006. 生长调节剂对落叶松杂种生根和幼苗生长的影响. 北京林业大学学报, 28(2): 68-72. |
孙晓梅, 张守攻, 周德义, 等. 2008. 落叶松种间及种内和种间杂种家系间物候变异与早期选择. 林业科学, 44(1): 77-84. |
王战, 张颂云. 1992. 中国落叶松林. 北京: 中国林业出版社.
|
王景章, 从培艳. 1980. 落叶松杂交育种及F1性状遗传. 林业科学, 16(1): 49-52. |
王笑山, 马浩, 王建华, 等. 2000. 落叶松杂种大规模繁殖配套技术研究. 林业科学研究, 13(5): 469-476. DOI:10.3321/j.issn:1001-1498.2000.05.003 |
王笑山, 郑先武, 王建华, 等. 1995. 采穗园母株生根性状遗传变异和选择效应. 林业科学研究, 8(1): 48-53. |
杨书文, 鞠永贵, 张世英, 等. 1985. 落叶松杂种优势的研究. 东北林业大学学报, 13(1): 30-36. |
张含国, 袁桂华, 李希才, 等. 1998. 落叶松生长和材性杂种优势的研究. 东北林业大学学报, 26(3): 25-28. |
张含国, 张成林, 兰士波, 等. 2005. 落叶松杂种优势分析及家系选择. 南京林业大学学报:自然科学版, 29(3): 69-72. |
Baltunis B S, Greenwood M S, Eysteinsson T. 1998. Hybrid vigor in Larix: growth of intra- and interspecific hybrids of Larix decidua, L. kaempferi after 5 years. Silvae Genet, 47: 288-293. |
Farmer J R, Durst J T, Shaotang D, et al. 1992. Effect of clones, primary ramets, and age of stock plants on tamarack rooting. Silvae Genetica, 41(1): 22-24. |
John A. 1979. Propagation of hybrid larch by summer and winter cuttings. Silvae Genetica, 28(5/6): 220-225. |
Kyle P, Michael G. 1997. Mass production of hybrid larch families//Annual Report and Research Summary of the Cooperative Forestry Research Unit. University of Maine, Orono, Maine 04469: Miscellaneous Report 406, 24-26.
|
Li B L, Wyckoff G W. 1994. Breeding strategies for Larix decidua, L. leptolepis and their hybrids in the United States. For Genet, 1: 65-72. |
Luc E P. 2004. Roles of European and Japanese larch in the genetic control of growth, architecture and wood quality traits in interspecific hybrids (Larix × eurolepis Henry). Ann For Sci, 61: 25-33. DOI:10.1051/forest:2003081 |
Macdonald J, Wood R F, Edwards M V, et al. 1957. Exotic forest trees in Great Britain//Forestry Commission Bulletin 30. London: HMSO.
|
Mason W L. 1989. Vegetative propagation of hybrid larch (Larix × eurolepis Henry) using winter cuttings. Forestry Supplement, 62: 189-199. |
Matyssek R, Schulze E D. 1987. Heterosis in hybrid larch (Larix decidua × leptolepis)Ⅱ: Growth characteristics. Trees, 1: 225-231. DOI:10.1007/BF01816820 |
Svensson J C, Mckeand S E, Allen H L, et al. 1999. Genetic variation in height and volume of Loblolly Pine open-pollinated families during canopy closure. Silvae Genetica, 48(3/4): 204-208. |