 2008, Vol. 44
2008, Vol. 44文章信息
- 杨春花, 周小平, 王小明.
- Yang Chunhua, Zhou Xiaoping, Wang Xiaoming.
- 卧龙自然保护区华西箭竹地上生物量回归模型
- Regression Models of Above-Ground Biomass of Fargesia nitida in Dengsheng Area in Wolong Nature Reserve
- 林业科学, 2008, 44(3): 113-123.
- Scientia Silvae Sinicae, 2008, 44(3): 113-123.
-
文章历史
- 收稿日期:2006-08-21
-
作者相关文章



2. 卧龙自然保护区管理局 中国保护大熊猫研究中心 汶川 623006
2. China Conservation and Research Center for Giant Panda, Wolong Nature Reserve Wenchuan 623006
生物量作为生态系统研究的本底数据, 是评价植物生长状况和生态系统功能的重要指标。群落多样性与生态系统生产力之间的关系是当今生物多样性研究领域中的焦点之一, 地上生物量是初级生产力的重要组成部分和表现形式(李凯辉等, 2007)。研究表明, 地上生物量与多样性指数存在不同形式的相关关系(Guo et al., 1998; 施济普等, 2001; 李凯辉等, 2007)。
中国素有“竹子王国"之称, 全国竹类植物共有48个属, 500多种(陈绪和, 2004)。竹子具有广泛的经济用途, 如可用于造纸的大木竹(Bambusa wenchouensis)、笋用的雷竹(Phyllostachys praecox)、做竹器的台湾桂竹(Phyllostachys makinoi)等, 同时又具有重要的生态价值, 如可用于水土保持的绿竹(Dendrocalamus oldhami)、植被恢复的芦竹(Aundp donax)等。研究竹子的生物量, 有利于评价经营措施、发展丰产技术、制定合理的经营开发措施, 有利于推进防护林体系建设。
有些竹种类如冷箭竹(Bashania fangiana)是一些珍稀、濒危的野生动物如大熊猫(Ailuropoda melanoleuca)、小熊猫(Ailurus fulgens)、毛冠鹿(Elaphodus cephalophus)的主要食物,研究这类竹子的地上部分生物量是野生动物管理的基本方面。由于食竹野生动物如大熊猫、小熊猫在一年中的不同季节, 选食竹子的不同部位(地上), 因此, 其主食竹生物量在不同器官中的分配特征, 在很大程度上反映了竹林的树冠结构、生产潜力和生活周期(吴福忠等, 2005), 关系到这些野生动物的生存。
生物量的传统研究方法有二氧化碳平衡法、微气象场法和收获法(薛立等, 2004)。竹子生物量的测定一般用收获法, 如皆伐法(王太鑫等, 2005)、标准地和平均木法(余英等, 2005), 但是出于操作方便的考虑, 最常用的是回归法。竹子的基径增长属于竹材的质量生长, 是竹株经过不断吸收养分而使细胞内贮藏的物质不断增多, 幼嫩组织不断木质化、纤维化, 导致竹株质量不断增加所形成的。竹子的高度主要取决于立地条件, 竹子所处的立地条件越好, 竹子吸收土壤养分的速度、效率就越高, 竹高直接影响着竹子各器官生物量的大小(郑容妹等, 2003)。因此, 用基径、高度等因子可以预测竹子的生物量。迄今, 已有10多种竹子的生物量回归模型被研究、报道, 如缺苞箭竹(Fargesia denudata)、苦竹(Pleioblastus amarus)、邛竹(Qiongzhuea tumidinoda)等(刘兴良等, 1994; 郑容妹等, 2003; 董文渊等, 2002; 林新春等, 2004)。
华西箭竹(Fargesia nitida)为箭竹属长三角鞘系、多年生一次性开花结实、秆丛生或近散生的木本克隆植物, 高2~5 m, 粗1~2 cm,在我国主要产于甘肃南部和四川西部, 为大熊猫的主食竹之一(秦自生等, 1993; 宋利霞等, 2006)。如在黄龙各种箭竹中, 以华西箭竹的分布最广(胡杰等, 2000); 在陕西周至老县城自然保护区, 华西箭竹成为仅次于秦岭箭竹的第2大竹种(冯永辉等, 2006)。同时, 华西箭竹为丛生竹(极少数为散生), 低矮密生, 姿态优美, 开发森林旅游时可作为园林景观。
关于华西箭竹的研究, 主要集中在:不同亚高山暗针叶林林冠环境(林下、中林窗、大林窗和林缘旷地)下华西箭竹分株种群的结构特征(宋利霞等, 2006)、无性系数量特征、无性系根茎特征、分株生物量以及分株形态特征对比研究(陶建平等, 2006); 华西箭竹对亚高山森林不同演替阶段(阔叶林、针阔混交林与针叶林)物种多样性与乔木更新的影响(王永健等, 2007); 华西箭竹对岷江冷杉主要乔木树种幼苗结构及分布格局(王微等, 2007)、幼龄植株种群结构动态(李媛等, 2007a)和幼苗更新(李媛等, 2007b)的影响; 以及华西箭竹种群与大熊猫种群的数学模型(程国忠等, 2001)、大熊猫对华西箭竹选择与利用的研究(胡杰等, 2000)等。
研究华西箭竹地上部分生物量, 可以了解其蓄积量, 并可据此推算它对以其为食的各种野生动物如大熊猫、小熊猫等的承载量, 还可以评价森林生态系统状况。但对华西箭竹的生物量回归模型研究至今尚无报道, 本研究拟在这方面进行探讨, 研究成果可为自然保护区资源管理、野生动物保护提供基础资料和参考。
1 地点与时间研究在卧龙自然保护区邓生保护站野牛沟区域进行。该地区为四川盆地向青藏高原的过渡地带, 属于青藏高原气候区, 研究区域内竹种以华西箭竹的分布较多, 为林下灌木层优势种(秦自生等, 1993;宋利霞等, 2006)。
采集华西箭竹标准竹的区域中心设置的大样方(20 m×20 m)位于102°59′ E、30°52′ N, 海拔约2 750 m, 坡向280°(西坡), 坡度约15°, 为岷江冷杉原始暗针叶林。华西箭竹生物量调查设置的2条样线位于野牛沟两侧, 最高海拔3 500 m, 最低2 740 m, 以白桦(Betula platyphylla)、红桦(Betula albo-sinensis)、房线槭(Acer franchetii)-岷江冷杉(Abies faxoniana)针阔混交原始林为主, 平均坡度25°左右, 平均植被盖度85%以上, 有竹地占70%左右, 有竹地平均竹枝叶盖度约60%。
标准竹的测量、烘干在2005年12月进行。生物量野外调查的时间是2005年5月、2005年12月和2006年5月。
2 研究方法 2.1 标准竹的测量方法1) 选竹 以所设大样方为中心并开始取样, 向前、后每隔50 m有代表性地选取当年生、2年生、多年生华西箭竹各5株, 共5个取样点, 取样区带跨度为200 m, 共选取75株标准竹(每龄级各25株, 均为完整未断尖长势正常竹), 齐地面砍断。
2) 测量 用电子游标卡尺(精度为0.02 mm)、米尺(精确到0.1 cm)、天平(精确到0.1 g)分别测量基径、杆高度、单株质量(下同)。之后分离杆、枝、叶各部分, 并分别测其质量。
3) 取样 杆、枝、叶分年龄分别取样, 其中枝剪短至6~8 cm, 杆剪短至5~8 cm并劈开成宽度不超过1 cm的长条。
4) 烘干 用恒温烘箱, 先在65 ℃烘至恒重(杆和枝24 h, 叶15 h), 称量, 然后升温至105 ℃烘至恒重(杆和枝3 h, 叶2 h), 称量。
2.2 竹生物量的调查方法1) 样线的设置 沿野牛沟两侧设置大致平行的A、B 2条样线, 样线距沟>40 m。
2) 样方的设置 ① 2005年冬:在样线上每隔200 m设置一个20 m×20 m的大样方, 共36个大样方, 其中有竹样方26个。每个大样方又分成4个10 m×10 m的中样方, 目估法测量每个中样方的竹盖度(根覆盖度)。又于每个中样方满布竹根处有代表性地各选取一个1 m×1 m的小样方, 计数每个小样方中的当年生、2年生、多年生竹株数, 每个小样方中有代表性地选取6株竹(每龄级2株), 测其基径和高度。并对其中1个小样方用绳子固定, 所测量的6株竹也绑好标记, 便于复查。② 2005年春:与2005年冬之不同在于, 每个小样方中测量8株竹, 不区分竹龄。③ 2006年春:复查2005年冬样方。不同之处:只设1个小样方, 即2005年冬作了标记的小样方, 计数其中竹数, 测量作了标记的6株竹的基径和高度。
3) 竹子的称量和取样 2005年冬, 于每个有竹大样方有代表性地选取华西箭竹15株(每龄级5株), 齐地砍断, 用电子吊钩称(精度为5 g)称每龄级5株总质量。每样线有代表性地选取每龄级全竹各1~2株, 剪断、劈细,用以测干物质。
4) 生物量的计算公式 某大样方每平方米华西箭竹生物量(g)为: 2005年春
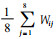
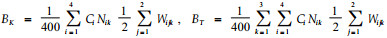
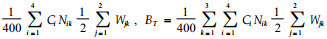
整个研究区域有竹地华西箭竹每平方米生物量为26个有竹大样方生物量的均值, 即: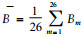
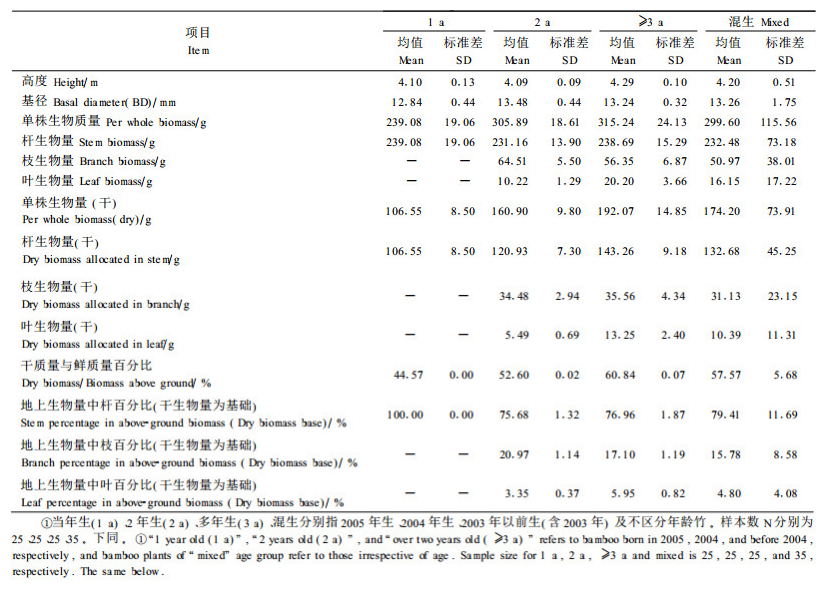
75株标准竹的描述性统计结果见表 1。其中, 当年生竹还没有生出枝和叶。
|
|
做单株生物量及单株各地上器官生物量关于基径或高度的散点图(图 1)。由图可以看出, 各龄级单株总生物量、单株杆生物量以及单株枝生物量与基径和高度之间, 均有明显的正相关关系。单株干生物量与2预报因子的回归趋势与鲜质量的相似, 限于篇幅, 仅给出鲜生物的散点图。

|
图 1 华西箭竹地上生物量与高度及与基径的散点图 Figure 1 Scatterplots of above-ground biomass with height and with basal diameter of Fargesia nitida A:单株地上生物量Per whole above-ground biomass;B:单株杆生物量Per whole biomass allocated in stem;C:单株枝生物量Per whole biomass allocated in branch;D:单株叶生物量Per whole biomass allocated in leaf. |
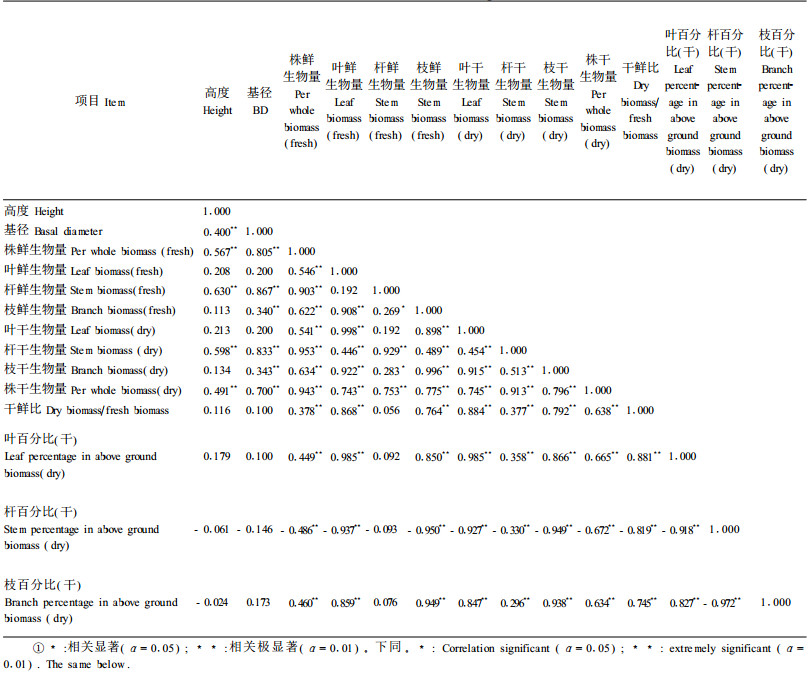
各变量值之间的Spearman相关系数见表 2。可见, 大部分项目均与基径和高度呈极显著相关, 因此, 可用基径、高度作为预报因子预测单株生物量等变量。而各地上构件器官生物量百分比与基径和高度则无显著相关(α=0.05), 因此, 用基径、高度不能预测杆、枝、叶所占的比例, 同理, 也不能预测干生物量与鲜生物量的比例。
|
|
在正态分布检验的基础上, 采用t检验, 对各龄级两两之间进行多重比较(表 3)。选用Kruskal-Wallis H非参数检验法检验各龄级间的差异(表 3)。
|
|
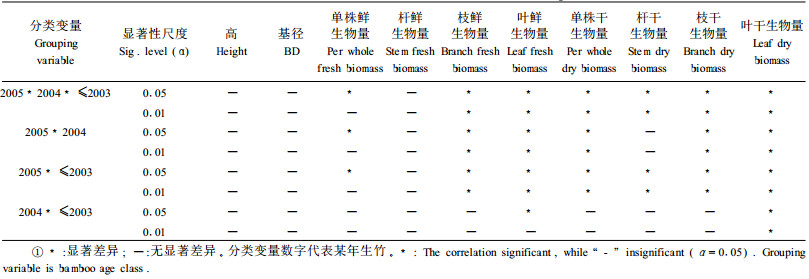
由表 3可见, 许多项目各龄级间有显著差异, 所以对各龄级华西箭竹的回归方程将分别推导和讨论。
3.1.5 生物量的回归模型(方程)基于75株标准竹的测量结果, 建立了各龄级华西箭竹地上生物量关于基径和高的一元及二元回归模型。
在2005年春的调查中, 未区分竹子年龄, 为比较该季节的生物量与后来2次调查的差异, 还建立了未区分年龄竹(“混竹")的生物量回归模型。根据26个有竹样方中1年生、2年生、多年生的竹子株数比例(据2005年冬调查结果)(考虑权重, 权重为各中样方的竹盖度)——10.90%:17.23%:71.87%, 分别从75株标准竹中(随机)选择1年生、2年生、多年生各4、6、25株, 组成混合样本, 建立了“混竹”生物量回归模型。
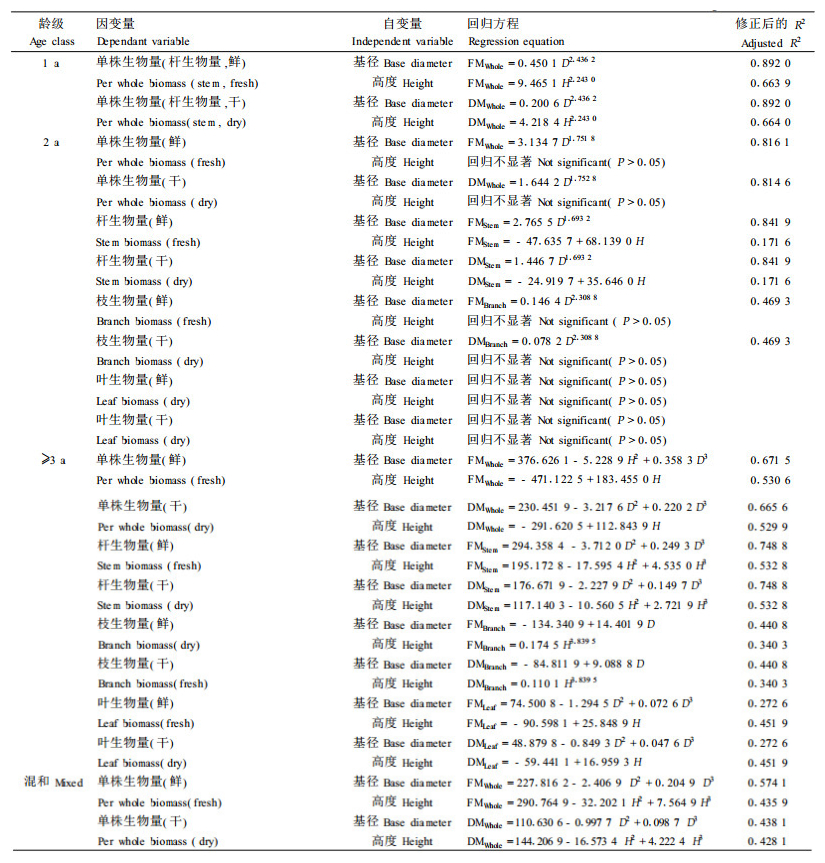
1) 一元回归方程应用线性、二次方、三次方、对数、倒数、逻辑斯蒂、生长指数、幂函数、指数等函数分别对各龄级华西箭竹的测量结果进行拟合。限于篇幅, 仅将调整后判定系数R2和回归效果显著性检验的F值最大的一种回归方程列出(表 4)。
|
|
选用一个自变量, 有可能会导致预报信息的丢失。因此, 有必要将基径和高度都纳入回归方程。采用逐步回归法(判据为:P<0.05, 自变量被引入模型; P>0.10, 自变量被剔除出模型)建立了生物量关于基径和高度的二元线性回归方程和二元幂函数回归方程(表 5)。
|
|
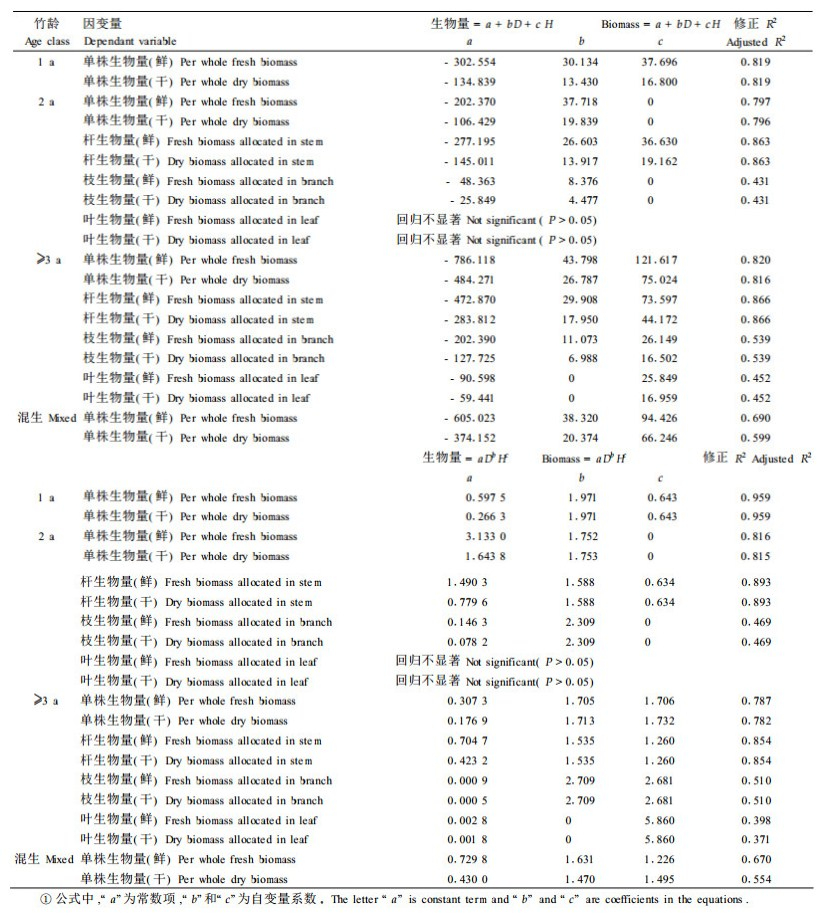
由表 5可见, 2种模型的预报精度差别不是很大(从修正R2判断)。
2种模型都不适合2年生叶生物量, 不过, 叶生物量可由株生物量-杆生物量-枝生物量来求算。
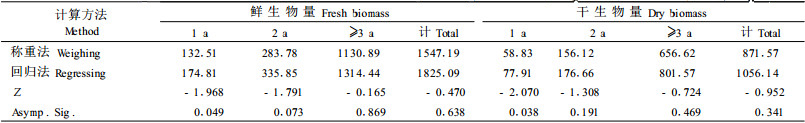
3.2 模型的检验(生物量2种求算方法——回归法和称重法的比较)对2005冬26个有竹样方的调查结果, 用二元幂函数回归法和收割称重法, 分别求出每单位面积生物量(表 6)。经检验, 在α=0.05下, 用2种方法所计算的生物量之间除1年生外, 均无显著差异; 在α=0.01下, 则均无显著差异(表 6)。说明所建立的模型可以较准确地预测华西箭竹的地上生物量。
|
|
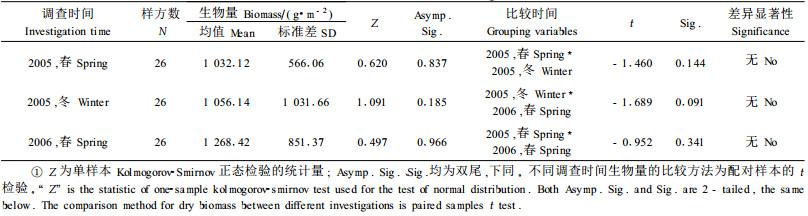
应用表 5的二元幂函数回归模型及野外调查数据, 计算出了邓生野牛沟区域26个有竹样方不同时间华西箭竹地上生物量, 并进行了比较。
3.3.1 2005年春、2005年冬、2006年春华西箭竹地上生物量比较经配对样本t检验, 不同调查时间的生物量(干物质基础)两两之间均无显著差异(α=0.05)(表 7)。
|
|
经非参检验, 2季节间各龄级地上总生物量(干物质基础)均不显著(α=0.05) (表 8)。在α=0.05下, 2005年冬、2006年春各龄级华西箭竹各单位面积生物量(干)除多年生杆外, 均无显著差异; 在α=0.01下, 则各项均无显著差异(表 8)。
|
|

陶建平等(2006)曾采用平均木法研究了同一地区(邓生)华西箭竹的生物量。方法是:在亚高山暗针叶林4种不同林冠环境中各设8个4 m×4 m样方, 在每一林冠环境下随机测量20株华西箭竹(没区分年龄), 冲洗烘干后称量各部分质量, 以20株的平均值代表某林冠环境华西箭竹无性系分株(各构件)的生物量。从本研究结果来看, 回归法和称重法算得的生物量除当年生外无显著差异(表 6), 但是, 回归法具有操作简便、受外界干扰小等优点, 所以回归法优于称重法。同时, 竹类是不同年龄立竹构成的异龄林, 由于生长年限的不同, 各年龄的竹株所进行的光合作用的时间有所差异, 因此其生物量及其分配必然存在差异(林益明等, 1998), 生物量在不同年龄竹的分配是不同的(郑郁善等, 1998; 薛萍等, 2006), 笔者的统计检验也证明了这一点(表 3), 因此, 不同龄级竹的生物量应分别求算、讨论。此外, 陶建平等(2006)的研究表明, 不同林冠环境间, 分株生物量、分株构件生物量和分株构件生物量分配百分率均有显著差异, 开敞的林冠环境有利于华西箭竹的生长和生物量积累。但是, 林冠环境的差异是否造成了单株生物量积累和分配的不同, 即:不同林冠环境下华西箭竹的单株地上生物量及单株地上构件器官生物量回归方程系数是否相同,此有待验证。
相同立竹度情况下, 林分立竹的径级分配也是影响竹林生物量的主要因子之一(陈双林等, 2004), 异速生长模型仅适用于特定的基径、高度范围(Alvaro et al., 2006)。本研究中未区分标准竹的径级、高度级, 主要是基于2点考虑:第一, 华西箭竹是一种小径竹, 基径和高度范围都比较小, 种群的基径和高度变异范围较小; 第二, 如果为每一径级或每一高度级的华西箭竹分别建立一套回归模型, 那么当把如此众多的模型应用于研究实践时, 势必会带来许多繁琐, 因而并不实用。
单木生物量模型误差的来源主要有因子选择和模型采用的数学形式2个方面(郭清文等, 2006)。由于竹生长参数的选择受竹种、年龄、器官的影响, 因此, 如何依据其生物学特性选择适当的回归参数(因子)是提高生物量回归法测定精度的关键(郑郁善等, 1998)。从以往的研究看, 多数研究选择的因子都集中在竹径(基径、胸径)和竹高(全高、枝下高)等2方面参数。从表 4可知, 除2年生叶和多年生叶外, 各龄级华西箭竹地上生物量及地上各构件器官生物量与基径的回归程度都明显高于与高度的回归程度, 这一点与文献上许多竹种的研究报道是一致的,这也是有的研究者对有些竹种仅采用竹径作为预报因子的原因(郑容妹等, 2003; 刘兴良等; 1994; 董文渊, 2002)。华西箭竹地上生物量与胸径、与枝下高的回归关系如何尚有待进一步研究。
竹生物量与基径和高度的回归模型多以幂函数为最佳, 如石竹(Phyllostachy nuda)、苦竹等(黄宗安, 2000; 林新春等, 2004)。从表 4可见, 华西箭竹当年生竹、混生竹, 以及2年生除叶质量(不能由基径或高度预报)和杆重与高度的回归关系外, 各项一元回归模型均以幂函数回归最为显著。但是, 多年生竹一元回归模型却多以三次方最佳。今后有必要增大标准竹取样量, 进一步验证、研判华西箭竹的生物量回归模型。
从图 1可见, 相同高度下, 单株生物量和单株枝生物量均是1年生最小, 多年生次之, 2年生最大; 这和与其同域分布的冷箭竹和拐棍竹的情况不同, 后两者是1年生最小, 2年生次之, 多年生最大。这是由于相同高度下, 2年生竹长势相对粗壮(基径较大)(表 1), 而多年生竹是3年及3年以上各年龄的混和竹, 基株年龄已达约23年(陶建平等, 2006), 老龄竹自身活力降低, 缺枝少叶, 生物量积累弱。此外, 与冷箭竹和拐棍竹相比, 华西箭竹的叶比例较低(表 1)(杨春花,2007), 这一方面是由于所调查竹种竹龄偏高, 密度过大(调查过程中也发现, 死竹比例相当高), 叶生长空间受限制; 另一方面, 是由于华西箭竹在亚高山地区广布于森林林窗、林下和林缘所构成的异质生境中(陶建平等, 2006), 其所生长环境中光资源(光照强度)优于冷箭竹(Bashania fangiana)和拐棍竹(Fargesia robusta), 喜荫的华西箭竹便采取减小叶面积、叶生物量来减少水分的蒸发。
本研究与以往研究的一个最大不同点在于, 竹子林分生物量的调查范围和取样力度较大。笔者的调查充分考虑到华西箭竹分布环境的异质性, 每隔一定距离(200 m)机械性设置一个大样方, 每个大样方又分4个中样方(调查竹盖度)和4个小样方(调查竹密度)并有代表性地选取竹子进行检尺, 并且考虑权重(权重为各中样方的竹盖度), 兼顾了代表性和准确性, 因而所求算的林分生物量更可信。
对于散生竹, 估计大样方竹密度可用枝叶覆盖度也可用根盖度, 当然相应地, 小样方应在布满枝叶处及布满竹根处选取。华西箭竹为丛生竹(也有少数是散生的), 若在枝叶覆盖处选择小样方, 会导致较大误差, 因此估计大样方竹密度时宜用根覆盖度, 相应地, 应在满布竹根处选小样方。由于华西箭竹为丛生竹, 每蔟面积都较大(直径一般在4~5 m以上), 且竹根着生非常紧密, 使得簇中心竹在非破坏性取样前提下无法测到, 只能在簇外围测量, 这可能会导致野外调查时一定的误差。密集的大簇中央处华西箭竹生物量是否适合本文回归模型, 有待进一步验证。
2005年春到2005年冬, 中间经历了一次竹发笋的生物学过程, 但笔者调查所得到的2个季节的生物量却几近相等, 差异不显著, 说明动物消耗和自然死亡的竹生物量大体抵消了竹生长量, 使得生物量处于动态平衡之中。也有可能是2次调查的方法不完全一致所致。同样, 2006年春比2005年冬生物量略有增长(但差异不显著), 也可能是这2方面原因所致。
综上所述, 以基径和高度为自变量建立的地上生物量的一元及二元回归方程(尤其是二元幂函数回归方程)能很好地预测华西箭竹的地上生物量。本文所建立的华西箭竹地上生物量回归模型可在实践中应用。今后的研究可在不同林冠环境、不同径级、不同因子方面进一步验证、研判华西箭竹的生物量回归模型。
陈双林, 吴柏林, 吴明, 等. 2004. 退化低丘红壤新造毛竹林地上部分生物量的研究. 江西农业大学学报, 26(4): 527-531. DOI:10.3969/j.issn.1000-2286.2004.04.011 |
陈绪和. 2004. 发展竹业促进扶贫和经济发展. 世界竹藤通讯, 2(2): 41-44. DOI:10.3969/j.issn.1672-0431.2004.02.008 |
程国忠, 张洪德, 胡杰, 等. 2001. 黄龙寺自然保护区大熊猫与华西箭竹两种群的数学模型. 四川师范学院学报:自然科学版, 22(2): 106-111. |
董文渊, 黄宝龙, 谢泽轩, 等. 2002. 筇竹无性系种群生物量结构与动态研究. 林业科学研究, 15(4): 416-420. DOI:10.3321/j.issn:1001-1498.2002.04.009 |
冯永辉, 冯鲁田, 雍严格, 等. 2006. 秦岭大熊猫主食竹的分类学研究(Ⅱ). 西北大学学报:自然科学版, (36): 101-102. |
郭清文, 冯仲科, 张彦林, 等. 2006. 单木生物量模型误差分析与定权方法探讨. 中南林业调查规划, 25(1): 5-9. DOI:10.3969/j.issn.1003-6075.2006.01.002 |
胡杰, 胡锦矗, 屈植彪, 等. 2000. 黄龙大熊猫对华西箭竹选择与利用的研究. 动物学研究, 21(1): 48-51. DOI:10.3321/j.issn:0254-5853.2000.01.010 |
黄宗安. 2000. 石竹各器官生物量回归模型研究. 竹子研究汇刊, 19(4): 54-57. DOI:10.3969/j.issn.1000-6567.2000.04.010 |
李凯辉, 胡玉昆, 阿德力·麦地, 等. 2007. 天山南坡高寒草地物种多样性及地上生物量研究. 干旱区资源与环境, 21(1): 155-159. DOI:10.3969/j.issn.1003-7578.2007.01.032 |
李媛, 陶建平, 王永健, 等. 2007a. 暗针叶林下华西箭竹(Fargesia nitida)对岷江冷杉(Abies faxoniana)幼龄植株种群动态的影响. 生态学报, 27(3): 1041-1049. |
李媛, 陶建平, 王永健. 2007b. 亚高山暗针叶林林缘华西箭竹对岷江冷杉幼苗更新的影响. 植物生态学报, 31(2): 283-290. |
林新春, 方伟, 俞建新, 等. 2004. 苦竹各器官生物量模型. 浙江林学院学报, 21(2): 168-171. DOI:10.3969/j.issn.2095-0756.2004.02.011 |
林益明, 林鹏, 叶勇. 1998. 绿竹种群生物量结构研究. 竹子研究汇刊, 17(2): 9-13. |
刘兴良, 向性明. 1994. 王朗林区人工缺苞箭竹亚层数量特征及其模型. 林业科学, 30(5): 471-477. |
秦自生, 艾伦·泰勒, 蔡绪慎. 1993. 卧龙大熊猫生态环境的竹子与森林动态演替. 北京: 中国林业出版社, 13, 37-39; 52-57; 149-156; 170-210; 245-318.
|
施济普, 张光明, 白坤甲, 等. 2001. 黄竹侵入对群落生物量及植物多样性的影响. 植物资源与环境学报, 10(4): 34-37. DOI:10.3969/j.issn.1674-7895.2001.04.008 |
宋利霞, 陶建平, 王微, 等. 2006. 卧龙亚高山暗针叶林不同林冠环境下华西箭竹分株种群结构特征. 生态学报, 26(3): 730-736. DOI:10.3321/j.issn:1000-0933.2006.03.013 |
陶建平, 宋利霞. 2006. 亚高山暗针叶林不同林冠环境下华西箭竹的克隆可塑性. 生态学报, 26(12): 4019-4026. DOI:10.3321/j.issn:1000-0933.2006.12.013 |
王太鑫, 丁雨龙, 李继清, 等. 2005. 巴山木竹种群生物量结构研究. 竹子研究汇刊, 24(1): 19-24. DOI:10.3969/j.issn.1000-6567.2005.01.004 |
王微, 陶建平, 胡凯, 等. 2007. 华西箭竹对岷江冷杉林主要乔木树种幼苗结构及分布格局的影响. 林业科学, 43(1): 1-7. |
王永健, 陶建平, 李媛, 等. 2007. 华西箭竹对卧龙亚高山森林不同演替阶段物种多样性与乔木更新的影响. 林业科学, 43(2): 1-7. |
吴福忠, 王开运, 杨万勤, 等. 2005. 大熊猫主食竹群落系统生态学过程研究进展. 世界科技研究与发展, 27(3): 79-84. DOI:10.3969/j.issn.1006-6055.2005.03.015 |
徐道旺, 陈少红, 杨金满. 2004. 毛环竹笋用林生物量结构调查分析. 福建林业科技, 31(1): 67-70. DOI:10.3969/j.issn.1002-7351.2004.01.018 |
薛立, 杨鹏. 2004. 森林生物量研究综述. 福建林学院学报, 24(3): 283-288. DOI:10.3969/j.issn.1001-389X.2004.03.021 |
薛萍, 汪晓萍, 李锡泉. 2006. 湖南衡阳紫色页岩地区芦竹生物量的动态规律. 中南林学院学报, 26(3): 11-14. DOI:10.3969/j.issn.1673-923X.2006.03.002 |
杨春花.2007.放归大熊猫(Ailuropoda melanoleuca)预选栖息地评估——以卧龙为例.华东师范大学博士学位论文. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y1073646
|
余英, 费世民, 何亚平, 等. 2005. 长宁苦竹种群结构和地上生物量研究. 四川林业科技, 26(4): 90-93. DOI:10.3969/j.issn.1003-5508.2005.04.017 |
郑容妹, 郑郁善, 丁闽锋, 等. 2003. 苦竹生物量模型的研究. 福建林学院学报, 23(1): 61-64. DOI:10.3969/j.issn.1001-389X.2003.01.015 |
郑郁善, 洪伟. 1998. 毛竹丰产年龄结构模型与应用的研究. 林业科学, 34(3): 325-333. DOI:10.3321/j.issn:1001-1498.1998.03.017 |
Alvaro R, Florencia M. 2006. Growth, productivity, aboveground biomass, and carbon sequestration of pure and mixed native tree plantations in the Caribbean lowlands of Costa Rica. For Ecol Manage, 232: 168-178. DOI:10.1016/j.foreco.2006.05.067 |
Guo Q F, Berry W L. 1998. Species richness and biomass: dissection of the hump-shaped relationships. Eccology, 79(7): 2555-2559. DOI:10.1890/0012-9658(1998)079[2555:SRABDO]2.0.CO;2 |