 2008, Vol. 44
2008, Vol. 44文章信息
- 叶金山, 崔永兰, 杨文萍.
- Ye Jinshan, Cui Yonglan, Yang Wenping.
- 泡桐自然接干性状的遗传变异
- Genetic Variation of Paulownia's Natural Stem-Join Characters
- 林业科学, 2008, 44(3): 87-93.
- Scientia Silvae Sinicae, 2008, 44(3): 87-93.
-
文章历史
- 收稿日期:2005-10-31
- 修回日期:2006-11-23
-
作者相关文章


2. 上海师范大学生命科学学院 上海 2000234
2. College of life of Shanghai Normal University Shanghai 200234
由于泡桐的自然接干深刻影响着泡桐树干的高度、通直度、圆满度和材积并直接关系到桐材的利用范围和经济价值,因而泡桐的自然接干性问题始终受到人们的高度关注。翔实的史料表明,有关泡桐人工接干和干形培育的研究非常深入,其悠久的研究史甚至可以上溯到北宋陈翥的《桐谱》,而有关泡桐自然接干的研究却很薄弱(河南省《泡桐》编写组,1978;中国林业科学研究院泡桐组等,1982;倪善庆,1986;蒋建平,1990;蒋建平等,2000;熊耀国,1995;李占民,1996;王新建等,1999;侯元凯等,1999a;范国强等,2000;茹广欣等,2001)。这种状况自20世纪90年代后期以来有了明显改变。侯元凯等(1999b; 2000; 2001)以兰考泡桐(Paulownia elongata)为材料研究了泡桐不定芽的自然接干规律、苗木顶芽越冬特性和苗木顶芽水分变化规律。刘震等(2004;2005a; 2005b)以优良无性系"毛白33"为材料研究了泡桐顶侧芽休眠发育的温度特性、侧芽萌发成枝接干规律和下侧芽萌发成枝与上侧芽萌发接干间关系。崔永兰等(2001)研究了泡桐接干类型划分和干形相关性状。迄今未见有关泡桐自然接干性状遗传变异性的研究报道。本文从林木遗传育种学角度研究了泡桐自然接干性状的遗传变异规律,希望为认识和改良泡桐的自然接干性提供有用信息。
1 材料与方法 1.1 材料1) 泡桐属主要的种和变种的天然林和天然散生木:我国南、北桐区的河南、河北、北京、山东、安徽、江苏、江西、浙江、湖南、湖北、广东和福建等地自然接干状态下的泡桐天然林和天然散生木;2)杂种试验林:由泡桐属10个种和变种之间的正交、反交、三交和回交68个杂交组合的600余株杂交种单株组成,其杂交亲本种和变种是:白花泡桐(P. fortuneii)、毛泡桐(P. tomentosa)、兰考泡桐、南方泡桐(P. australis)、台湾泡桐(P. kawakamii)、川泡桐(P. fargesii)、鄂川泡桐(P. albiphloea)、楸叶泡桐(P. catalpifolia)、山明泡桐(P. lamprophylla)和成都泡桐(P. albiphloea var. chengtuensis);3)毛泡桐种源试验林:由来自全国各地的120个种源的毛泡桐组成;4)无性系试验林:由23个杂交种无性系和8个亲本种无性系共31个无性系组成。
以上后3类试验林地势平坦而彼此相连,位于豫东平原黄泛区腹地,约为34°20′ N,114°28′ E,海拔60 m,年均降水量678.5 mm, 年均气温14.4 ℃。林地土壤为黄河冲积土中的沙壤土,pH 7.0,地下水位3.5 m,肥力中等。3类试验林的树龄为12年,均按5 m×10 m株行距定植。其中无性系试验林为随机区组设计,31个无性系6株小区4次重复并统一规定以C125作为对照。
1.2 方法1) 野外观察:1998—2004年野外观察我国南北桐区泡桐天然林、天然散生木和3种试验林在野外自然条件和人工栽培条件下的自然接干性;2)性状测定:采用每木测量计算无性系试验林的22个自然接干性状:树高、主干高、接干高、全干高、胸径、主干基径、主干1/2径、主干端径、接干1/2径、主干材积、接干材积、全干材积、主干材积/全干材积、接干材积/全干材积、主干形数、接干形数、主干形率、接干形率、主干平均削度、通直度(L)、冠幅、丛枝病等级,采用4级通直度分类标准;3)统计分析:按中央直径法计算主干材积和接干材积,并令主干材积+接干材积=全干材积;以东西、南北冠幅的平均值代表冠幅性状值;以胸径为比较直径计算主干形率和接干形率;以胸高断面积和全干高为标准计算主干形数和接干形数;用长度除两头直径之差法计算各段树干的平均削度;按随机区组固定模型进行以单株性状值和以小区平均数为单位的一元和多元方差分析;多重比较用Duncan法;无性系选择用多维空间En多向量理论;同时还进行了主成分分析和相关分析。按林标(LYB)108-61原木材积表、国标(GB)198-63原条材积表和林标(LYB)104-60杉原木材积表所用公式计算与泡桐不同树干区分段同粗等长的原木和原条材积。
各性状的数据统计按完全随机区组模型进行分析,其线性统计模型为:
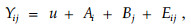
|
其中:γij为j区组i无性系的性状测定值;u为总体平均值;Ai为i无性系效应;Bj为j区组(重复)效应;Eij为机误。
无性系试验林的所有统计分析均采用SAS软件。本文省略了一些统计分析表格。
2 结果与分析 2.1 自然接干性状的遗传变异表 1给出了无性系试验林中31个无性系22个自然接干性状的平均数、标准差和变异系数。表 1数据表明自然接干性状在无性系水平上存在着广泛和明显的遗传变异性。按表 1中的性状变异系数CV的大小可将22个自然接干性状的遗传变异性划分为以下3种类型。
|
|
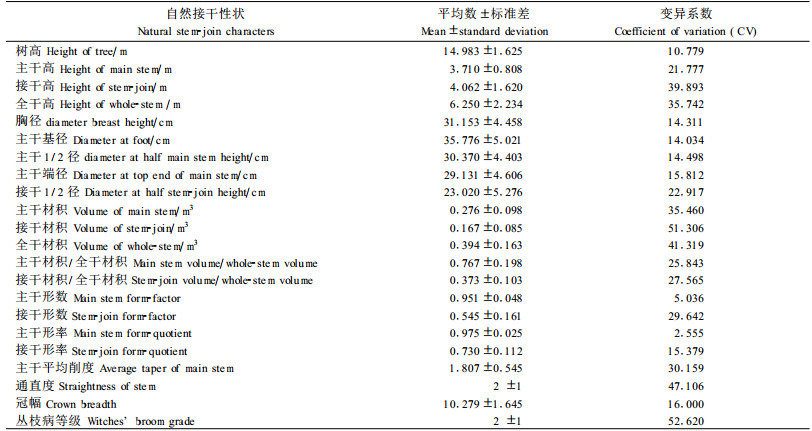
1) 高度变异型(CV≥30):有8个自然接干性状,即接干高、全干高、主干材积、接干材积、全干材积、主干平均削度、通直度、丛枝病等级的变异系数CV≥30而属于高度变异型,其CV变幅为30.159~52.620。
2) 中度变异型(15≤CV<30):有8个自然接干性状,即主干高、主干端径、接干1/2径、主干材积/全干材积、接干材积/全干材积、接干形数、接干形率、冠幅的变异系数属于中度变异型,其CV变幅为15.379~29.642。
3) 低度变异型(CV<15):有6个自然接干性状,即树高、胸径、主干基径、主干1/2径、主干形数、主干形率的变异系数属于低度变异型,其CV变幅为2.555~14.498。
仔细分析上述8个高度变异型自然接干性状的组成和关系可以看出:接干高性状在影响和决定其他自然接干性状的总体表现和最终状况上起着重要作用。其原因在于:1)接干高性状的变异直接影响了全干高、接干材积、接干材积/全干材积、接干形数和接干形率的变异;2)接干高性状正是通过着生于其上的枝叶的光合作用,间接影响了除通直度和丛枝病等级之外的、所有其他自然接干性状的生长发育状况和变异性表现(因通直度和丛枝病等级分别直接受控于接干侧芽的向上生长竞争性和泡桐丛枝病抗性基因,与光合作用关系不密切)。
由于泡桐是自然整枝作用非常强烈的强阳性速生树种,因此包括全部的种、变种、变型和杂交种在内的所有泡桐属树种的植株都普遍呈现出这样一种共同的树冠结构特征,即泡桐植株的枝叶除了在其个体生命史的最初1~3 a着生于主干外,总是着生在接干上,并随着接干的生长而不断上移。由于光合作用是所有自养有机体一切有机物质和能量的最终来源与产量形成的基础,因此泡桐的接干高性状正是通过着生于其上的枝叶的光合作用而对其他自然接干性状产生直接和间接影响,而其他自然接干性状也必须依靠接干高性状才能存在和发展。因此接干高性状不但在研究泡桐自然接干性状的遗传变异性上起着重要作用,而且也在泡桐自然接干性状的遗传改良中占据着关键地位。人们衡量和评价用材树种树干特征的最基本标准就是要求其树干必须通直圆满。仅就通常情况而言,绝大多数用材树种的树干一般总是圆满的,但并非总是通直的。这条基本林学原理普遍适用于包括泡桐在内的所有用材树种。泡桐树高生长上独特的接干和干形问题都是由泡桐特有的假二叉分枝现象造成并直接损害了泡桐树干的通直度,而研究和解决泡桐自然接干性问题的关键和难点最终都可归结到泡桐通直度性状上的L=1和L≠1的遗传变异性上。毫无疑问,只要消除了L≠1的情形,泡桐的自然接干效果就与其他始终以顶芽进行树高生长的乔木用材树种的树高生长和干形特征没有本质差别了,而只要实现了L=1就意味着实现了泡桐自然接干性状遗传改良的最终目标。因此富有成效的泡桐自然接干性状的遗传改良工作必须高度重视和深入研究通直度性状遗传变异的基本规律和表现形式。
2.2 自然接干类型划分关于泡桐自然接干类型划分问题,早期的研究者提出了连续接干型、间歇接干型和不易接干型3种划分法(中国林业科学研究院泡桐组等,1982;倪善庆,1986)。后来崔永兰等(2001)又将泡桐的自然接干类型划分为连续接干、徒长枝接干和难接干3种类型。
我们根据对泡桐属的主要种、变种和无性系在自然生长条件下(即材料部分中的第1类天然林和天然散生木)和人工栽培条件下(即材料部分中的第2、3和4类试验林)的自然接干类型的观察结果初步提出了以下“三分法”标准以划分泡桐的自然接干类型。
1) 按是否逐年都接干的特性而划分为连续接干型和间断接干型。前者是每年都接干而后者是间断接干。泡桐属中除兰考泡桐的自然接干生长表现出"长长停停"的特性而属于间断接干型外,其余的泡桐种和杂交无性系的自然接干都属于连续接干型。
2) 按接干侧芽的部位高低而划分为顶端侧芽接干型和内部侧芽接干型。前者由树干最上端侧芽接干而后者由树干内部侧芽接干。白花泡桐、楸叶泡桐、毛泡桐、台湾泡桐和杂交无性系多为顶端侧芽接干型,而兰考泡桐、川泡桐和南方泡桐则多为内部侧芽接干型。由于内部侧芽接干型的泡桐在栽植后2~4年容易从树冠内部萌生徒长枝,并由徒长枝形成接干,因而这种接干类型也称为徒长枝接干型。
3) 按接干的理想程度而划分为优良接干型、中间接干型和不良接干型。一般而言,前者的自然接干性强而能形成高而通直圆满的全干;后者自然接干性差而使全干非常低矮,中者的自然接干性中等而通常形成主干和接干的分段连接或粗细不均匀连接,并在桐材利用上产生了按主干材和接干材分段造材利用的现象。泡桐属中属于前者的有白花泡桐和楸叶泡桐,属于中者的有兰考泡桐、南方泡桐和川泡桐,而属于后者的有毛泡桐和台湾泡桐。
上述“三分法”中的前2种属于自然分类(根据接干侧芽的生物学表现)而后1种则体现了人为分类的思想(根据用材树种用于板材、家具材和胶合板材时的造材要求),因此“三分法”既明确了3种划分标准之间的逻辑关系(完全独立和平行的划分关系),又有利于消除观察角度和评价标准上的分歧和混乱现象。
2.3 不同自然接干类型的接干优劣性评价虽然从泡桐树高生长的不同方式和实现途径的角度无法比较不同的自然接干类型, 但是从泡桐木材主要用于板材、家具材和胶合板材的用途和价值的角度却又是能够比较和必须比较其不同自然接干类型。由于在除纸浆材和薪炭材之外的所有泡桐用材林都无一例外地要求泡桐树干长得高而通直圆满和削度小,而泡桐遗传改良追求的最基本目标就是要为泡桐的用材林选育出自然接干性状表现特别优良的无性系,因此从用材树种的树干特性来评价泡桐的不同自然接干类型的接干效果和接干优劣性无疑具有理论研究和指导生产的双重意义。
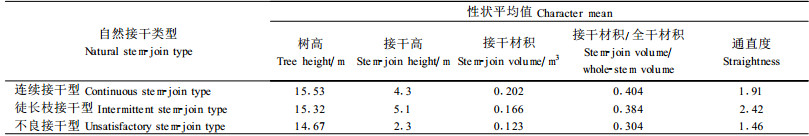
表 2、3证明了接干高和通直度的平均性状值在不同接干类型间存在着较大差异。虽然平均树高在3种自然接干类型中基本相同,但是连续接干型和徒长枝接干型的平均接干高明显高于不良接干型的平均接干高,而徒长枝接干型的平均接干高又大于连续接干型的平均接干高。表 2、3的数据和野外观察表明有些无性系完全可以凭借其极好的自然接干性而使全干长得高而通直圆满,从而达到桐材利用上的理想干形要求。由于接干高和通直度直接影响了接干材积和全干材积,因而接干高和通直度在不同自然接干类型间的差异性直接导致不同类型间的接干材积和接干材积/全干材积比的明显差异。此外,表 2数据还清楚表明虽然连续接干型的平均接干高小于徒长枝接干型的平均接干高,但是其平均接干材积却占徒长枝接干型平均接干材积的121.7%,从而导致其接干材积/全干材积比也高于徒长枝接干型的接干材积/全干材积。造成这种现象的原因是徒长枝接干型的树高生长呈现间断接干和接干位置不确定,因而降低了接干的粗度生长并导致了接干与主干的粗细不均匀连接或错位连接,最终损害了全干的通直度、圆满度、材积和利用价值。由此可知3种自然接干类型中当以连续接干型最好,并表明在泡桐遗传育种和造林上应该强调培育和推广属于连续自然接干型1级通直度标准的速生优质抗病新品种。
|
|
|
|

一元方差分析结果证明:在31个无性系的22个自然接干性状中只有通直度性状没有显著性差异,其余21个自然接干性状都有极显著差异。
多元方差分析结果证明:自然接干性状在无性系间存在极显著差异。
Duncan分析表明:1)在杂交无性系间主干高、主干基径和冠幅性状无显著性差异;2)大多数杂交无性系的胸径性状与对照的差异没有达到显著水平;3)少数自然接干性状表现特别优良的速生杂交无性系如“9501"、“9502"和“8508-2"等虽然在树高、胸径、主干高、主干基径和冠幅性状上与对照的差异不显著,但在接干高、全干高、接干材积、全干材积、接干材积/全干材积、主干形数、主干形率、接干形数、接干形率、主干平均削度和通直度性状上均与对照存在显著差异;4)“9501"、“9502"和“8508-2"在绝大多数自然接干性状上无显著性差异。
主成分分析揭示第一主成分(贡献率50.80%)为接干材积/全干材积、接干材积、接干1/2径和接干高,第二主成分(贡献率18.00%)为主干材积,而第三主成分(贡献率6.91%)为主干平均削度。实际上,第一主成分是指接干材积因子,第二主成分是指主干材积因子,而第三主成分则清楚指出了主干平均削度在自然接干性状中的重要性。这3个主成分的累积贡献率已达75.91%。
相关分析发现泡桐自然接干性状间的相关性变化很大。一些重要性状间的相关性极强,例如接干高与全干高、接干1/2径、接干形数、接干材积和接干材积/全干材积都呈极显著的正相关,它们的相关系数高达0.88~0.94,接干1/2径与接干形数、接干材积和接干材积/全干材积也呈极显著的正相关,它们的相关系数高达0.89~0.96;有些性状间的相关性很低,例如主干形数与全干材积、主干平均削度与接干材积、树高与通直度的相关系数分别为0.11、0.04和0.14;有些性状间却呈极显著的负相关,例如主干高与主干形数、主干高与接干材积、接干高与主干材积的相关系数分别为-0.66、-0.47和-0.25。
表 4运用多维空间En多向量理论综合评定了31个无性系的接干材积、主干材积和主干平均削度,结果发现“9502"、“8508-2"和“9501"的Pi值最小,分别为0.29、0.38和0.39,因而是所有无性系中自然接干性表现最好的3个优良无性系。
|
|
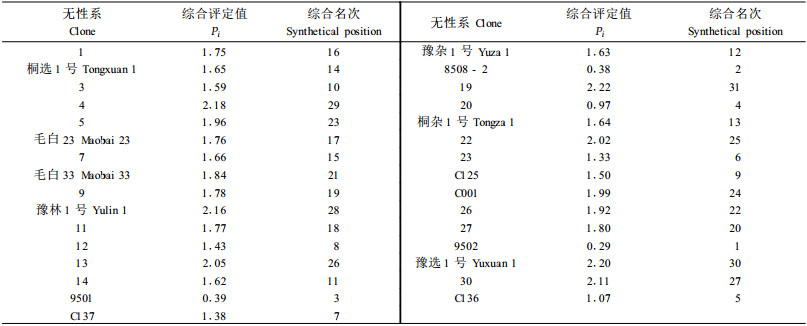
表 5估算了3个入选无性系“9501"、“9502"和“8508-2"的接干材积、主干材积和主干平均削度性状的平均值、广义遗传力和遗传增益。
|
|

表 6给出了入选无性系3株样木的31个自然接干性状值以从测树学上定量描述“9501"、“9502"和“8508-2"的优良自然接干性。从表 6可知“9501"、“9502"和“8508-2"的全干高分别高达10.7、9.5和12.0 m,其无节良材高度都超过6 m;接干高度分别高达7.2、4.5和9.0 m;全干通直度都为1级且主干形数分别高达0.87、0.93和0.98。表 6中不同树干段的平均削度值也证明了这3个无性系的树干具有极好的圆满度。此外,从材积比较角度将泡桐主干原木和全干原条的树干材积与我国通用原木和原条材积表中同粗等长的树干材积相比较无疑能更加具体和形象地描述上述3个优良无性系的树干圆满度:与同粗等长的所有树种的原条材积相比,“9501"、“9502"和“8508-2"的原条材积分别高达84.7%、107.3%和126.8%;与同粗等长的除杉木以外的所有树种的原木材积相比,“9501"、“9502"和“8508-2"的原木材积分别高达82.8%、99.8%和89.6%;与同粗等长的杉原木材积相比,“9501"、“9502"和“8508-2"的主干原木的材积分别高达84.1%、97.2%和92.3%。
|
|
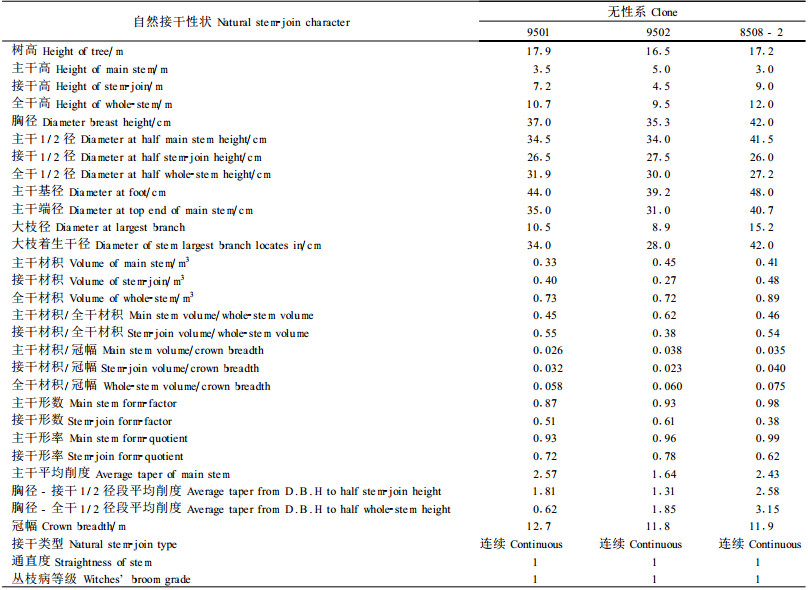
泡桐的自然接干性状在无性系水平上存在着广泛和明显的遗传变异性。其中有8个自然接干性状(接干高、全干高、主干材积、接干材积、全干材积、主干平均削度、通直度和丛枝病等级)属于高度变异型(CV>30),其CV变幅为30.159~52.620。接干高性状在影响和决定其他自然接干性状的总体表现和最终状况上起着重要作用,而通直度性状在泡桐自然接干性状的遗传改良中起着至关重要的作用。
初步提出了划分泡桐自然接干类型的“三分法”标准,并证明了连续接干型是最好的自然接干形式。
一元方差分析证明:在31个无性系的22个自然接干性状中只有通直度性状没有显著性差异,其余21个自然接干性状都有极显著差异。多元方差分析证明:自然接干性状在无性系间存在极显著差异。Duncan分析表明:大多数杂交无性系的胸径性状与对照的差异不显著;少数自然接干性状表现特别优良的速生杂交无性系如“9501"、“9502"和“8508-2"等虽然在树高、胸径、主干高、主干基径和冠幅性状上与对照的差异不显著,但在其他自然接干性状上均与对照存在显著差异;“9501"、“9502"和“8508-2"在绝大多数自然接干性状上无显著性差异。主成分分析揭示接干材积因子、主干材积因子和主干平均削度分别为第一、第二和第三主成分,三者的累积贡献率达75.91%。相关分析发现泡桐自然接干性状间的相关性变化非常大。有些性状间呈极显著的正相关,有些性状间的相关性很低,还有一些性状间却呈极显著的负相关性。多维空间En多向量理论评定出“9501"、“9502"和“8508-2"是所有无性系中自然接干性表现最好的3个优良无性系。简单估算了“9501"、“9502"和“8508-2"接干材积、主干材积和主干平均削度的平均值、广义遗传力和遗传增益。从入选无性系样木的31个自然接干性状值和同粗等长的原木和原条材积比较角度具体形象地描述了入选无性系的优良自然接干性。
整个研究工作最终说明了泡桐自然接干性状在无性系水平上存在着广泛和明显的遗传变异,很多自然接干性状的遗传变异具有重要的育种学价值并能进行遗传改良,而利用常规杂交育种技术就可以有效地改良这些自然接干性状,从而培育出自然接干性状表现十分优良的速生优质抗病泡桐新品种。
崔永兰, 吕国政, 石俊阁. 2001. 泡桐接干类型划分及干形相关性状的分析. 河南农业大学学报, 35(1): 70-73. DOI:10.3969/j.issn.1000-2340.2001.01.020 |
范国强, 王安亭, 王国周, 等. 2000. 接干和施肥对不同初植苗高泡桐幼树主干生长影响的研究. 林业科学研究, 13(6): 628-633. DOI:10.3321/j.issn:1001-1498.2000.06.009 |
河南省《泡桐》编写组. 1978. 泡桐. 北京: 科学出版社.
|
侯元凯, 翟明普. 1999a. 泡桐干形培育研究进展. 林业科学, 35(3): 76-83. |
侯元凯, 翟明普, 娄季松, 等. 1999b. 兰考泡桐不定芽自然接干规律研究. 北京林业大学学报, 21(3): 14-19. |
侯元凯, 翟明普, 张俊昌, 等. 2000. 兰考泡桐苗木顶芽越冬特性初步研究. 河南农业大学学报, 34(2): 196-200. DOI:10.3969/j.issn.1000-2340.2000.02.028 |
侯元凯, 翟明普, 聂爱社, 等. 2001. 兰考泡桐苗木顶芽水分变化规律研究. 北京林业大学学报, 23(6): 17-21. DOI:10.3321/j.issn:1000-1522.2001.06.005 |
蒋建平. 1990. 泡桐栽培学. 北京: 中国林业出版社, 263-279.
|
蒋建平, 范国强, 李培玉, 等. 2000. 泡桐主干与树冠生长相关关系的研究. 河南农业大学学报, 34(2): 127-129. DOI:10.3969/j.issn.1000-2340.2000.02.007 |
李占民. 1996. 泡桐单芽接干试验研究. 河北林业科技, (1): 11-13. |
刘震, 何松林, 王艳梅, 等. 2004. 泡桐顶侧芽休眠发育的温度特性研究. 林业科学, 40(3): 46-50. |
刘震, 毕会涛, 蒋建平, 等. 2005a. 泡桐侧芽萌发成枝接干规律. 林业科学, 41(4): 42-47. |
刘震, 耿晓东, 秦素玲, 等. 2005b. 泡桐下侧芽萌发成枝与上侧芽萌发接干间关系. 北京林业大学学报, 27(5): 65-69. |
倪善庆. 1986. 泡桐. 南京: 江苏科学技术出版社.
|
茹广欣, 袁金玲, 冯胜, 等. 2001. 泡桐不同无性系生长性状及抗病性分析. 河南农业大学学报, 35(1): 53-56. DOI:10.3969/j.issn.1000-2340.2001.01.016 |
王新建, 张秋娟, 杨玉金, 等. 1999. 31个泡桐无性系性状相关性研究. 林业科技通讯, (2): 7-11. |
熊耀国. 1995. 泡桐遗传改良. 北京: 中国科学技术出版社, 45-133.
|
中国林业科学研究院泡桐组, 河南省商丘地区林业局. 1982. 泡桐研究. 北京: 中国林业出版社, 171-176.
|