 2008, Vol. 44
2008, Vol. 44文章信息
- 魏文毅, 王贵禧, 梁丽松, 樊金拴.
- Wei Wenyi, Wang Guixi, Liang Lisong, Fan Jinshuan.
- ‘八月脆’桃果实气调贮藏过程中品质及相关酶活性的变化
- Changes of Quality and Some Enzyme Activities of 'Bayuecui' Peach during Controlled Atmosphere Storage
- 林业科学, 2008, 44(3): 81-86.
- Scientia Silvae Sinicae, 2008, 44(3): 81-86.
-
文章历史
- 收稿日期:2007-05-16
-
作者相关文章



2. 黑龙江八一农垦大学 大庆 163319;
3. 中国林业科学研究院林业研究所 国家林业局林木培育实验室 北京 100091
2. HLJ August First Land Reclamation University Daqing 163319;
3. Research Institute of Forestry CAF Beijing 100091
桃是具有呼吸跃变的水果,采后在常温下极易软化腐烂,在低温下易发生冷害。受冷害后的桃果肉褐变,质地硬化、木质化或絮状败坏,后熟时不能正常软化,固有风味变淡甚至丧失,有的甚至产生异味或苦味(王友升等, 2003)。目前国内外有许多关于桃采后生理及贮藏保鲜技术的研究报道,大多致力于解决低温导致的冷害问题,气调贮藏是当今果蔬贮藏方式中被认为是效果最好的贮藏技术,它能有效地减轻果蔬的冷害。5%O2+10%CO2的气体条件下冬雪蜜桃贮藏82 d,果实不褐变,92 d时果实的风味仍然正常(田世平等, 2001)。采用(1~3)%O2+(3~8)%CO2的气调贮藏‘大久保’和‘绿化9号’,其贮藏期达到60 d以上,未发生衰败症状,最长贮藏期达到120 d(吕昌文等, 1993)。本研究以晚熟品种‘八月脆’桃(Amygdalus persica 'Bayuecui')为试材,研究桃果实在气调贮藏过程中果肉品质及相关生理生化指标的变化,同时研究在不同贮藏期出库后熟3 d(20 ℃)时的果实硬度变化及果肉的褐变情况,为桃的产业化气调贮藏提供理论与技术指导。
1 材料与方法 1.1 材料'八月脆'桃于2005年9月10日和2006年9月3日采自北京市平谷区,当天运至中国林业科学研究院林业研究所果品保鲜实验室,选择大小均匀、无伤害、商业成熟的果实进行有关处理,成熟度的划分参照SB/T 10090-92。2005年进行初步研究,2006年进一步重复试验,本文采用的是2006年的研究结果。
1.2 试验处理供试果实在0~1 ℃下预冷48 h,进行以下处理。1)对照:用打孔聚乙烯袋(0.045 mm)包装后于0~1 ℃恒温冷藏,用CK表示;2)气调贮藏(0~1 ℃,10%O2+10%CO2) (魏文毅, 2004),用CA表示。每处理重复3次。
气调贮藏果实密封于20 L的广口瓶中,每瓶装10 kg,通过流量计持续通入配制好的气体,流量约70~100 mL·min-1。定期取样,一部分立即进行有关指标的测定,另一部分置于20 ℃后熟3 d后对后熟期间的果肉硬度和褐变指标进行测定。
1.3 测定指标与方法1) 硬度 用硬度计(意大利产,FT327型,探头直径11 mm)测定果实的去皮硬度。可溶性固形物含量(SSC)用手持折光仪测定。膜渗透率用DDSJ-308型电导率仪测定,用相对电导率表示。
2) 褐变指数 褐变指数=Σ(褐变级数×果实个数)/(最高级数×果实总个数)×100。
0级:果肉无褐变;1级:果肉有轻度褐变;2级:果肉有明显褐变但褐变面积不及1/3;3级:褐变面积为1/3~2/3;4级:果肉褐变面积大于2/3。
3) 出汁率 称取20个果肉块(直径10 mm,厚3 mm),装入垫有吸水棉的离心管中,1 700×g离心10 min。以果肉块离心后的质量损失率作为出汁率(茅林春等, 1999)。
4) 多酚氧化酶(PPO)活性测定 提取液的制备:取10 g去皮果肉,加0.5 g PVPP于20mL 0.2 mol·L-1磷酸缓冲液(pH 6.4)中,冰浴研磨,4 ℃冰冻离心机12 000 rpm离心20 min,取上清液测定酶活性。
活性测定:PPO活性测定参照Galeazzi等(1981)的方法,并加以改进。将0.2 mL粗酶提取液加入3 mL 0.1 mol·L-1的邻苯二酚溶液(用0.2 mol·L-1 pH 6.4的磷酸缓冲液配成)中。反应温度为30 ℃,加酶液后5 s开始扫描20 s内398 nm处吸光值变化,以每分钟ΔOD398变化0.01表示一个酶活性单位(U):
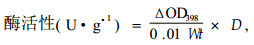
|
式中:ΔOD398为反应时间内吸光度的变化;W为样品鲜质量(g);t为反应时间(min);D为酶液总体积与测定时取用酶液体积的比值。
5) 过氧化物酶(POD)和过氧化氢酶(CAT)活性 分别采用愈创木酚法(李合生, 2000)、紫外吸收法(高俊风, 2000)。
6) 果胶甲酯酶(PE)(Buescher et al., 1978)和多聚半乳糖醛酸酶(PG)活性测定(Chan et al., 1981)。
7) 可溶性糖含量(蒽酮比色法)(李合生, 2000),果胶含量(咔唑比色法)(韩雅珊, 1992)。
1.4 数据差异性分析本试验数据用SPSS软件进行统计处理,采用ANOVA进行邓肯氏多重差异分析。
2 结果与分析 2.1 气调贮藏对果实硬度和可溶性糖含量的影响由图 1可知:CA贮藏果实硬度在第10 d和第20 d时的硬度显著高于冷藏对照(P<0.05),随后迅速下降到一稳定的水平;CK贮藏果的硬度在贮藏的前期和中期大致保持稳定。在30 d以后,CA与CK贮藏果的硬度值无显著差异(P<0.05),并且在近60 d的贮藏期内硬度均较高,这可能与‘八月脆’桃属于非溶质型桃品种有关。

|
图 1 气调贮藏对‘八月脆’桃硬度的影响 Figure 1 Effect of CA storage on firmness of 'Bayuecui' peach |
常温3 d后熟时,CA和CK中果实的硬度均显著低于刚出库时果实的硬度(图 1),但是在第30 d后熟时,CK中果实的硬度减少量逐渐降低,到贮藏第60 d时出现了明显的不软化现象,而CA中果实的硬度减少量则相对稳定。因为出库后不能正常软化是冷害的一种表现,说明CK中果实在第30 d时就已经受到了轻微冷害,随后冷害程度加深,而CA则有助于减轻冷害的发生。
从图 2分析得知,CA和CK贮藏果实的可溶性糖含量均呈下降趋势,但CA在第50 d以前下降非常缓慢,随后才快速下降,贮藏末期接近CK的含量, 并且CA贮藏果的可溶性糖含量在贮藏第20 d以后显著高于CK的同期值(P<0.05)。说明气调贮藏可使桃果实在低温逆境条件下,能够保持更多的可溶性糖,降低渗透势和冰点,从而适应或提高耐受低温的能力。

|
图 2 气调贮藏对‘八月脆’桃可溶性糖含量的影响 Figure 2 Effect of CA storage on soluble sugar content of 'Bayuecui' peach —◆— CK, —△— CA.下同The same below. |
贮藏果实的出汁率既与果实的冷害程度有关,又与果实的成熟软化程度有关。由图 3可知,CK贮藏果的出汁率在贮藏20 d(14.7%)后逐渐升高,到第30 d时达到18.0%,接着从第40 d的18.1%快速升高到贮藏结束时的25.1%;40 d以后出汁率的快速上升,与果实出库后不能正常软化的现象一致(图 1),说明是由于果实发生冷害有关。而CA贮藏果的出汁率在整个贮期内变化不大,且显著低于CK贮藏果(P<0.05)。

|
图 3 气调贮藏对‘八月脆’桃出汁率的影响 Figure 3 Effect of CA storage on juice extraction rate of 'Bayuecui' peach |
冷敏果实在长时间低温胁迫下,细胞膜的完整性受到破坏,细胞膜透性升高,离子渗出率增大。冷藏的桃果实组织的相对电导率随着贮期的延长大幅度升高,尤其是在贮藏30 d以后,一直处于较高的水平(图 4),说明普通冷藏在20 d以内对‘八月脆’桃果实的细胞结构破坏程度较低,而30 d后果实受到的冷害程度加深。气调贮藏桃果实的相对电导率在贮藏期间基本保持稳定,并且显著低于普通冷藏的(P<0.05)(图 4)。说明气调贮藏对于保持细胞膜的稳定性、防御冷害的发生有较好的效果。

|
图 4 气调贮藏对‘八月脆’桃膜透性的影响 Figure 4 Effect of CA storage on membrane permeability of 'Bayuecui' peach |
冷藏果实的出汁率与组织电导率骤然升高在时间上基本一致,分析表明显著相关(r=0.843 8)。
2.3 气调贮藏对果胶甲酯酶(PE)和多聚半乳糖醛酸酶(PG)活性的影响不同处理果实的PE活性表现出近似的变化趋势(图 5)。在冷藏条件下,处理与CK贮藏果的PE活性整体上呈增加趋势,并且均在第40 d时出现PE活性高峰,但在整个贮藏期间,处理与CK贮藏果的PE活性差异不显著(P>0.05)。说明气调贮藏对PE的活性没有直接的影响。

|
图 5 气调贮藏对‘八月脆’桃PE活性的影响 Figure 5 Effect of CA storage on the PE activity of 'Bayuecui' peach |
由图 6可知,在贮期的30 d前和30 d后,CA和CK贮藏果的PG活性具有显著差异(P<0.05),尤其在第50 d时CK贮藏果的PG活性明显高于CA。一方面说明CA对桃果实PG活性的影响具有阶段性,在贮藏前期对PG活性的提高有利于果胶物质的正常代谢,使果实对低温的适应性增强,而后期对PG活性的适当抑制可以减慢由于果胶质的快速降解导致的细胞降解和分离引起的过度软化。另一方面说明CK的高活性是底物诱导的结果,因为PG作用的底物是去甲酯化的果胶质(果胶酸),虽然处理与CK贮藏果的PE活性差异不显著,但在贮藏的后期CK贮藏果的PE活性值略高于CK(图 5)。

|
图 6 气调贮藏对‘八月脆’桃PG活性的影响 Figure 6 Effect of CA storage on the PG activity of 'Bayuecui' peach |
从图 7可知,在贮藏期间,CA和CK贮藏果实的POD活性总体呈下降趋势,CA贮藏果的POD活性在整个贮藏期间波动性较大,在贮藏初期20 d内大幅度下降,接着又快速上升到开始降低的水平,随后降低。CK贮藏果的POD活性在整个过程中较稳定,且显著高于CA(P<0.05)。POD是一种诱导酶,即在果实受到伤害或衰老时,体内积累的自由基含量增加,诱导POD的活性上升。本研究中,冷藏对照果的POD活性比CA贮藏果高,说明冷藏对照果受冷害或衰老的程度比气调果重。

|
图 7 气调贮藏对‘八月脆’桃的POD活性的影响 Figure 7 Effect of CA storage on the POD activity of 'Bayuecui' peach |
2种贮藏条件下桃果实的CAT活性变化如图 8所示,在整体上,两种条件下的桃果实CAT活性均呈升高的趋势,但CA贮藏果的CAT活性显著高于CK(P<0.05)。说明CA明显提高了‘八月脆’桃在冷藏期间的CAT活性,从而减轻了‘八月脆’桃果实在贮藏期间因过氧化问题而造成的伤害,尤其是对细胞膜的结构破坏,亦表明CAT在保持‘八月脆’桃果实组织细胞膜的完整性中具有相当重要的作用,这与前述内容一致。

|
图 8 气调贮藏对‘八月脆’桃CAT活性的影响 Figure 8 Effect of CA storage on the CAT activity of 'Bayuecui' peach |
如图 9可知,各处理果的PPO活性均呈降低趋势。CK贮藏果的PPO活性在贮藏20 d以前相对稳定(平均值50.63 U·g-1),到第30 d时已大幅度下降,直到贮藏结束时一直维持在下降后的低水平(平均值14.81 U·g-1)。CA贮藏果的PPO活性在贮藏30 d内大幅度降低(平均值38.88 U·g-1),随后略有上升,接着快速下降到最低点(18 U·g-1),末期回升到下降时的水平,但从第30 d后直至贮藏结束,CA处理的PPO活性变化相对稳定(平均27.21 U·g-1)。从整个贮藏过程来看,在贮藏30 d内CA贮藏果的PPO活性显著低于CK(P<0.05),而从第30 d到第60 d的贮期内则显著高于CK(P=0.05)。结果说明,气调贮藏在贮藏前期能够明显抑制‘八月脆’桃果实的PPO活性,后期则相反。

|
图 9 气调贮藏对‘八月脆’桃PPO活性的影响 Figure 9 Effect of CA storage on the PPO activity of 'Bayuecui' peach |
褐变统计情况表明(图 10):CA的果实在贮藏期内均无明显褐变,只是在第60 d时出现极轻微的褐变,褐变指数为10.0%;而CK在第50 d褐变指数为5.0%,到贮藏第60 d时褐变指数达到25.0%, 显著高于CA(P<0.05)。在3 d后熟过程中,CA在第40 d才开始出现褐变,褐变指数为10.0%,随后褐变程度缓慢上升,末期后熟时,CA仅为17.5%;而CK贮藏果实在第20 d后熟时就出现褐变(2.5%),以后褐变骤增,第30 d时褐变指数达到50%,末期后熟时尽达85.0%,同时伴随有严重的腐烂。总的说明CA不仅能延缓冷藏下和后熟时‘八月脆’桃褐变出现的时间(第10 d、第20 d),而且能明显降低果实的褐变程度,减少腐烂。另外,CA贮藏果实的果表颜色在2种过程中均保持采摘时的水平,十分艳丽,CK后熟果的颜色较暗淡。

|
图 10 气调贮藏对‘八月脆’桃褐变的影响 Figure 10 Effect of CA storage on flesh browning of 'Bayuecui' peach |
桃果实在低温环境中受到冷害后表现出汁液减少、粉质化或木质化、在常温后熟时不能正常软化和果肉褐变等(茅林春等, 2000a)。本试验中,在气调贮藏(10%O2, 10%CO2)前期‘八月脆’桃果实表现出硬度上升的现象(图 1),我们认为这不是冷害造成的,而是由于CA贮藏果为了对低温的适应,果胶质的代谢速率相对较低而出现的暂时“回生”现象,同样的现象在杏果实贮藏和中华寿桃气调贮藏(4%~6%O2,7%~9%CO2)过程中也出现过(Taylor et al., 1995;孙萍等, 2003),因此说明气调贮藏在控制冷害的同时更有利于‘八月脆’桃果实的正常成熟,从而保持了果实在贮藏后期的良好品质。
一些研究者认为,桃果实的出汁率可以用来反映桃果实絮败程度,同时可以作为判断桃果实冷害程度的一个理化指标(茅林春等, 1999;2000b;王阳光等, 2001),并且多数认为冷害果实的出汁率往往呈降低趋势。本研究中,普通冷藏果实的出汁率在贮藏20 d后呈急剧上升趋势(图 3),这与‘大久保’桃在冷害温度(5 ℃)贮藏下的出汁率变化一致(王贵禧等, 2005),说明一些桃果实受到冷害后,出汁率并不一定降低,笔者认为这与桃的品种特性有关,溶质型的桃受低温冷害后多出现汁液减少,果肉质地粉质或木质化,非溶质型的桃则易出现汁液剧增,果肉发绵等症状。同时我们对普通冷藏条件下桃果实的出汁率与组织相对电导率的对比分析发现,两者变化基本一致并显著相关(图 3、4),这种现象在‘大久保’桃果实采后常温(20 ℃)下的成熟、衰老过程中也存在(王友升等, 2003),表明果实组织细胞膜的完整性由于低温伤害后逐渐破坏是导致出汁率升高的一个重要原因。
桃果实在遭受低温冷害后易出现果肉的褐变是影响桃采后品质的极其重要的一方面,目前关于果蔬的褐变机理都建立在组织细胞膜的完整性这一共同的基础之上,并同时对一些重要酶的活性进行研究。有关果蔬褐变的酶研究最多的是多酚氧化酶(PPO),它能在有氧条件下,将酚类物质催化氧化为醌,醌通过聚合反应产生有色物质,在果蔬组织褐变起着重要的作用(杜晓东, 1998;皮钰珍等, 2001;叶梅, 2005)。本试验中,气调贮藏(CA)和普通冷藏(CK)的桃果实在贮藏过程中的PPO活(图 9)性前期较高(CK显著高于CA),后期则较低(CK显著低于CA),同时CA贮藏果无论是在贮期内还是在3 d后熟时,不仅出现褐变的时间晚于CK,而且褐变程度也明显低于CK,尤其是在贮藏后期的常温3 d后熟时差异更大(图 10), 但是PPO活性与2种贮藏条件下果实的褐变程度无明显的相关性。分析可能原因为:1) CK贮藏果在贮藏前期的高PPO活性诱导了果实后期的严重褐变,或者说PPO催化酚类物质的褐变具有滞后效应; 2)在贮藏后期,CK贮藏果的组织电导率一直处于高水平(图 4),细胞膜受到明显的破坏,更有利于酶与酚类底物的结合,此时较低活性的PPO也有可能催化酶促褐变的进行; 3) CA贮藏果在贮藏后期相对明显的高PPO活性,这和PPO的生理功能(如抗病性)有关(赵伶俐等, 2005),而没有导致冷藏和后熟果的明显褐变是由于膜的完整性并未受到明显破坏,底物和酶仍呈区域分布; 4)褐变并不完全是由PPO酶促反应引起的,可能还有其他氧化反应或衰老进程与褐变紧密相关(姜爱丽等, 2002).如本试验中,冷藏果的POD的活性显著高于气调贮藏(10%O2,10%CO2),因为POD除了有清除植物体内自由基而减轻伤害的作用外,还与果实的成熟、衰老和褐变密切相关(韩涛等, 2000;林河通等, 2002;詹嘉红, 2005)。
杜晓东. 1998. 中国梨果实褐变机理及控制途径(综述). 河北农业技术师范学院学报, 12(2): 54-58. |
高俊风. 2000. 植物生理学实验技术. 西安: 世界图书出版社, 198-199.
|
韩涛, 李丽萍. 2000. 果实和蔬菜中的过氧化物酶. 食品与发酵工业, 26(1): 69-73. DOI:10.3321/j.issn:0253-990X.2000.01.018 |
韩雅珊. 1992. 食品化学实验指导. 北京: 北京农业大学出版社.
|
姜爱丽, 田世平, 徐勇, 等. 2002. 不同气体成分对甜樱桃果实采后生理及品质的影响. 中国农业科学, 35(1): 79-84. DOI:10.3321/j.issn:0578-1752.2002.01.016 |
吕昌文, 修德仁, 齐灵. 1993. 桃的采后生理研究及探讨. 天津农业科学, (2): 14-16. |
林河通, 席玙芳, 陈绍军. 2002. 果实贮藏期间的酶促褐变. 福州大学学报, 30(增): 696-703. |
李合生. 2000. 植物生理生化试验原理和技术. 北京: 高等教育出版社, 124.
|
茅林春, 应铁进, 张上隆. 1999. 桃果实絮败与果胶质变化和细胞壁结构的关系. 植物生理学报, 25(2): 121-126. DOI:10.3321/j.issn:1671-3877.1999.02.004 |
茅林春, 张上隆. 2000a. 果胶酶与桃果实冷害的关系. 植物生理学通讯, 36(3): 266-271. |
茅林春, 王阳光, 张上隆. 2000b. 热处理减缓桃果实的采后冷害. 浙江大学学报, 26(2): 137-140. |
皮钰珍, 马岩松, 王善广, 等. 2001. 桃采后及贮藏生理研究进展. 果树学报, 18(1): 53-56. |
孙萍, 孙锐. 2003. 气调贮藏对中华寿桃果实品质的影响. 莱阳农学院学报, 20(3): 209-210. DOI:10.3969/j.issn.1674-148X.2003.03.017 |
田世平, 徐勇, 姜爱丽. 2001. 冬雪蜜桃在气调冷藏期间品质及相关酶活性的变化. 中国农业科学, 34(6): 656-661. DOI:10.3321/j.issn:0578-1752.2001.06.014 |
王贵禧, 王友升, 梁丽松. 2005. 不同贮藏温度模式下大久保桃果实冷害及其品质劣变研究. 林业科学研究, 18(2): 114-119. DOI:10.3321/j.issn:1001-1498.2005.02.002 |
魏文毅. 2004. '八月脆'桃保鲜过程中相关生理变化的研究.西北农林科技大学硕士论文. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y641538
|
王友升, 王贵禧. 2003. 冷害桃果实品质劣变及其控制措施. 林业科学研究, 16(4): 465-472. DOI:10.3321/j.issn:1001-1498.2003.04.016 |
王阳光, 陆胜明, 茅林春. 2001. 热处理对冷藏桃果实的贮藏效果和生理作用. 福建果树, (3): 1-4. DOI:10.3969/j.issn.1004-6089.2001.03.001 |
叶梅. 2005. 植物组织褐变的研究进展. 重庆工商大学学报:自然科学版, 22(4): 326-329. |
赵伶俐, 范崇辉, 葛红, 等. 2005. 植物多酚氧化酶及其活性特征的研究进展. 西北林学院学报, 20(3): 156-159. DOI:10.3969/j.issn.1001-7461.2005.03.042 |
詹嘉红. 2005. 橄榄果实中PPO和POD活性抑制研究. 亚洲热带植物科学, 34(4): 14-16. |
Buescher R W, Fnmnanski R J. 1978. Role of pectinesterase and polygalacturonase in the formation of woollimness in peaches. J Food Sci, 43: 264-266. DOI:10.1111/jfds.1978.43.issue-1 |
Chan H T, Tam S Y T. 1981. Papaya polygalacturonase and its role in thermally injured ripening fruit. J Food Sci, 46: 190-191. DOI:10.1111/jfds.1981.46.issue-1 |
Galeazzi M A M, Sgarbieri V, Costantinides S M. 1981. Isolation, purification and physiocochemical characterization of polyphenol oxidase from dwarf variety of banana (Musa cavendishii). J Food Sci, 46: 150-155. DOI:10.1111/jfds.1981.46.issue-1 |
Taylor M A, Rabe E, Dodd M C. 1995. Effect of harvest maturity on suhstances internal conductivity, solids and gel breakdown in cold stored 'Songold' plums. Postharvest Biol Technol, 5: 285-294. DOI:10.1016/0925-5214(94)00040-Y |