 2008, Vol. 44
2008, Vol. 44文章信息
- 宋新章, 张智婷, 肖文发, 李冬生, 张慧玲.
- Song Xinzhang, Zhang Zhiting, Xiao Wenfa, Li Dongsheng, Zhang Huiling.
- 长白山杨桦次生林采伐林隙幼苗更新动态
- Regeneration Dynamics of Logging Gaps in Populus davidiana-Betula platyphylla Secondary Forests in Changbai Mountain
- 林业科学, 2008, 44(3): 13-20.
- Scientia Silvae Sinicae, 2008, 44(3): 13-20.
-
文章历史
- 收稿日期:2007-01-12
-
作者相关文章



2. 河北农业大学林学院 保定 071001;
3. 中国林业科学研究院森林生态环境与保护研究所 国家林业局森林生态环境重点实验室 北京 100091;
4. 国家林业局调查规划设计院 北京 100714
2. College of Forestry, Agricultural University of Hebei Baoding 071001;
3. Institute of Forest Ecology, Environment and Protection, CAF Key Laboratory of Forest Ecology and Environment of State Forestry Administration Beijing 100091;
4. Academy of Forest Inventory Planning, State Forestry Administration Beijing 100714
林隙干扰对植被更新具有重要作用,是森林循环更新的一个重要阶段。林隙在森林结构、动态和多样性维持中起着重要作用,已成为当前森林生态学研究的最活跃领域之一(Sipe et al., 1995; Runkle, 1998; 臧润国等, 1998b; 2004; 梁晓东等, 2001)。目前,我国在林隙动态方面的研究尚属起步阶段,主要在温带阔叶红松林(吴刚, 1998; 臧润国等, 1998a; 1999; 于振良等, 2001; 张春雨等, 2006)、亚热带常绿阔叶林(熊小刚等, 2002; 安树青等, 1997; 贺金生等, 1999; 刘金福等, 2003)、西南亚高山针叶林(刘庆等, 2002;刘庆,2004a;2004b; 吴宁, 1999)、海南岛热带雨林(臧润国等, 2001;2002;2004)中进行了一些工作, 主要描述天然林受到自然干扰(如火灾、风倒、树倒)后形成的林隙及其更新状况,对其机理、机制性的问题尚缺乏深入探讨,缺乏系统性和整体性,而国外已从早期的林隙特征研究转向林隙内生理生态学及相关机制等的研究,因此我国同国际的同类研究相比还存在较大差距。同时,对人为干扰(如经营采伐)造成的林隙及其更新也少有报道(安树青等, 1997; 郝清玉等, 1998; 朱教君, 2002; 宋新章等, 2006;2007)。间伐产生的林隙一般是小尺度和短暂的(Spies et al., 1989),但小尺度干扰对森林结构的发展及林地状况等起了关键作用(Lertzman et al., 1996; Myers et al., 2000),已成为全球森林动态和天然更新研究的热点(Gray et al., 1996; Mclaren et al., 1996; Jennings et al., 1999; Zhu et al., 2003)。天然次生林已成为我国森林资源的主体(朱教君, 2002),鉴于次生林在我国森林资源和生态环境建设中的重要地位和作用,进行次生林林隙更新动态方面的研究,探讨次生林演替的内在生态学机理,对于实现次生林的可持续经营具有重要的理论意义和应用价值。本文对长白山杨桦次生林采伐林隙内的更新状况进行了调查研究,旨在为探讨次生林的可持续经营与管理提供科学依据。
1 研究地概况研究在吉林省白河林业局杨桦次生林经营采伐迹地进行(42°05′—42°28′N,128°02′—128°64′E)。该区属温带季风影响下的大陆性季风气候,年均气温7.3~4.9 ℃,年降水量600~900 mm,全年日照时数2 271~2 503 h,全年无霜期109~141 d。海拔700 m,坡度1°~5°, 为比较平缓的平地与较浅的沟谷相交错的地形。土壤为火山灰形成的典型暗棕壤,土层厚达100 cm以上。林龄50年,属中龄林,伐后林分密度为556株·hm-2,林分蓄积量110 m3·hm-2,平均直径16 cm,平均树高15 m,郁闭度0.6。主要树种有山杨(Populus davidiana)、白桦(Betula platyphylla)、色木槭(Acer mono)、胡桃楸(Juglans mandshurica)、山荆(Malus baccata)、椴木(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、山槐(Maackia amurensis)、蒙古栎(Quercus mongolica)和春榆(Ulmus japonica)等, 主要下木有忍冬(Lonicera ruprechtiana)、暴马丁香(Syringa reticulate)、卫矛(Euonymus alatus)、榛子(Corylus heterophylla)等。主要地被物为莎草(Cyperus rotundus),盖度30%,分布均匀。采伐方式为择伐,采伐强度25%,择伐后形成了大小不一的林隙,采伐年度分别为2002年和1996年。
2 研究方法 2.1 调查方法调查于2005年7月进行。采用样线调查法(Runkle, 1982; 臧润国, 1998a;1999; 陈迪马等, 2005),将杨桦次生林2002年采伐迹地内某一随机点作为调查起点,沿罗盘仪所指方向,由北向南穿行,遇到林隙则停下来,调查冠林隙(center of gap, 简称CG)和扩展林隙(explanded gap, ,简称EG)的长和宽、形状、年龄、方向,记录林隙内伐根的数量和种类,记录林隙边缘立木的种类、树高、胸径、年龄等。在每个扩展林隙内设9个2 m×2 m研究样方,第一个样方设在林隙中心(用CC表示,下同),其余8个分别沿长短轴方向设在距离扩展林隙边缘的一半处和扩展林隙边缘处(分别用NN、SS、EE、WW表示林隙北部、南部、东部和西部边缘的样方,用CN、CS、CE、CW表示林隙中心至林隙北、南、东、西缘一半处的样方,下同)。具体样方设置如图 1所示(以椭圆形为例)。在每个样方内调查树高<1.5 m的更新幼苗和树高>1.5 m但胸径<4 cm的更新幼树的种类、数量、年龄、树高、基径等。在距林隙边缘10 m处的非林隙林分中设1个10 m×10 m的对照样地(林隙间相距较近时,则共用一个对照样地),调查样地内立木的种类、年龄、树高、胸径等,在每个对照样地内沿对角线方向机械设置9个2 m×2 m的小样方,记录更新幼苗、幼树的种类、数量、年龄、高度、胸径或基径等。用LAI-2000冠层分析仪观测各小样方内更新幼苗的叶面积指数,用9个样方叶面积指数的平均值作为该林隙或对照地更新幼苗的叶面积指数进行分析。用这种方法对18个采伐林隙和9个对照样地内的更新幼苗进行了调查。用同样的方法在1996年采伐迹地上调查了10个采伐林隙和6个对照样地。

|
图 1 样方设置示意图 Figure 1 Location of 9 research plots in gap GC:林隙中心Gap center;NN:扩展林隙北部林缘处North edge of expanded canopy;CN:林隙中心至林隙北部林缘距离的一半处Center of gap center to north edge;CS:林隙中心至南部林缘距离的一半处Center of gap center to south edge;SS:扩展林隙南部林缘处South edge of expanded canopy;EE:扩展林隙东部林缘处East edge of expanded canopy;CE:林隙中心至东部林缘距离的一半处Center of gap center to east edge;CW:林隙中心至西部林缘距离的一半处Center of gap center to west edge;WW:扩展林隙西部林缘处West edge of expanded canopy.下同The same below. |
林隙面积计算:本文采用冠林隙进行计算分析。所调查林隙大多呈椭圆形,故采用常用的椭圆形面积计算公式:A=πLW/4。式中:A为林隙面积(m2);L为冠林隙长度(m),W为冠林隙宽度(m)(胡理乐等, 2007)。
根据林隙面积的大小,以50 m2为一级对调查的林隙进行分级研究。由于林隙均是在2002年或1996年择伐时形成,至调查时已分别达到3年和9年,故将采伐林隙分为3龄林隙和9龄林隙。
3 结果与分析 3.1 采伐林隙的基本特征林隙中<50 m2的林隙占7%,50~100 m2的林隙占21%,100~150 m 2的林隙占36%,150~200 m2的林隙占29%,>200 m2的林隙占7%。可见采伐林隙面积以50~200 m2居多。3龄林隙占64.3%,9龄林隙占35.7%。28.6%的林隙呈圆形,71.4%的林隙呈椭圆形。若以长轴表示林隙方向,则椭圆形林隙中55%为南-北方向,30%为东北-西南方向,15%为东南-西北方向。冠林隙(CG)的长轴平均为(15.97±5.98) m,短轴平均为(8.66±3.82) m,长/短轴的比例平均为1.92;扩展林隙(EG)的长轴平均为(23.71±4.07) m,短轴平均为(14.66±3.62) m,长/短轴的比例平均为1.67。由此可见,长轴与短轴的比例变化较大,说明椭圆的曲率随林隙大小的变化有明显的变化。林隙一般由7~10株采伐木形成,采伐木主要由山杨、白桦和山槐组成,这可能与这些树种生长迅速,比其他树种较早成材而易遭到砍伐有关。
3.2 采伐林隙内树木的种类组成和数量特征为便于研究,分别从3龄和9龄不同大小级林隙中各选出2个代表性林隙,共16个林隙(其中<50 m2的林隙3龄和9龄各1个),对林隙内更新幼苗的状况进行了分析。结果表明,3龄林隙内的更新幼苗中,共有乔木12种,其中色木槭、山槐、花楸(Sorbus pohuashanensis)和山荆的个体数量较多(表 1),这4类树种占乔木总种数的33.23%,但却占到乔木个体总数的56%;共有灌木11种,其中卫矛、暴马丁香和忍冬个体数量较多,它们占灌木总种数的27.27%,却占灌木个体总数的82.81%(表 1)。在9龄林隙内的更新幼苗中,共有乔木13种,其中水曲柳、山荆、色木槭和假色槭(Acer pseudo-sieboldianum)个体数量较多,它们占乔木总种数的26.68%,但却占到个体总数的73.34%;共有灌木11种,其中暴马丁香、榛子和卫矛3树种占灌木总种数的21.42%,却占到灌木个体总数的85.35%。对照林分的更新幼苗中有乔木10种,灌木11种,其中乔木以水曲柳和山荆、灌木以卫矛和暴马丁香的个体数量最多,分别占个体数量的56.77%和58.99%。可见,在林隙形成初期,更新幼苗的种类组成并不十分丰富,而且乔、灌木的个体数量分布都极不均匀,种类间数量差别很大。同时,比较3龄和9龄的幼苗组成可以看出,随着林隙年龄的增加,新出现了一些更新树种如假色槭,一些树种所占个体数量的比例有较大提高,如水曲柳和山荆,而一些树种所占个体比例有所下降,如色木槭、花楸等。这是由于随着林隙形成时间的延长,光照条件发生了改变,林隙内的微生境也相应发生了变化,为具有不同生物学特性的树种提供了异质性的环境资源,一些不能适应这种变化的树种的个体数量逐渐下降,能够适应这种变化的树种的个体数量则逐渐上升。对比林隙内外的更新状况可以看出,林隙内更新的乔、灌木树种都比郁闭林分丰富,并且随着林隙年龄的增加,这一特点愈加显著。一些树种在林隙内外所占个体的比例差异较大,表明这类树种对林隙干扰的反应比较强烈,如色木槭、花楸、山槐、暴马丁香等树种适合于林隙更新,而水曲柳、山荆、卫矛等树种则更适合于在郁闭林下更新。
|
|
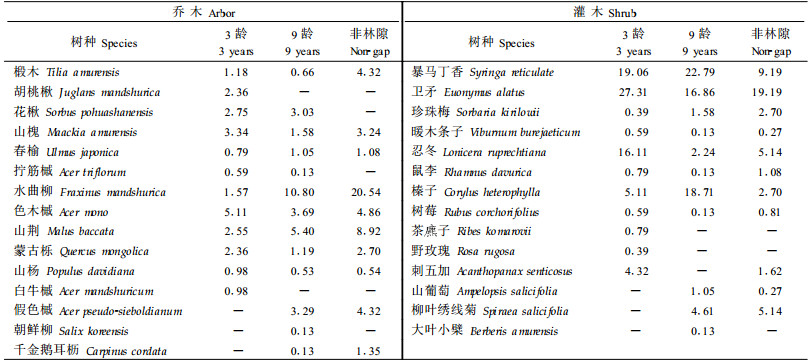
图 2表明,在3龄林隙内,更新幼苗的个体密度在50~100 m2大小的林隙中显著高于其他林隙;平均高和平均基径的变化趋势十分相近,在100~200 m2大小的林隙中较高;叶面积指数LAI在50~150 m2大小的林隙中较高。可以看出,LAI除与个体密度密切相关外,还与幼苗的平均高和平均基径有较大关系。种密度则随林隙面积的增加而变化较大,在>200 m2林隙中最低,在50~150 m2林隙和对照林分中最高,在其他林隙中基本一致。总起来看,<50 m2的林隙更新特征与对照林分相近。100~200 m2大小的林隙内更新幼苗的LAI、个体密度、平均高和平均基径均较高,表明该类大小的林隙尤其是100~150 m2大小的林隙较适合幼苗更新。上述4个特征指标均在>200 m2的林隙中最低,甚至低于对照林分,表明采伐林隙面积过大时其更新状况要差于中等面积大小的林隙,9龄林隙内的更新特征与3龄林隙相似,只是LAI、个体密度、平均高和平均基径的最大值均出现在100~150 m2林隙内,进一步说明该类大小的林隙有利于幼苗更新。

|
图 2 不同年龄不同大小林隙更新幼苗的特征(n=16) Figure 2 Trait of seedlings in gaps of different sizes and different ages |
出现这种现象的原因可能是:杨桦次生林林分密度不大,林冠稀疏,原本就存在许多小的林冠缝隙,所以小规模的采伐干扰(<50 m2)对林分环境的影响不大,故林隙内的更新与对照林无明显差异。随着采伐干扰程度加大,较多的大树被移出,大树根系对水分、养分的争夺随之减弱,林隙内的光照大量增加,资源有效性增大,更新环境明显改善,大大促进了幼苗的萌生与繁殖,不但个体密度迅速增加,反映幼苗生长状况的LAI、平均高和平均基径也显著提高。但当采伐干扰规模过大(>200 m2)时,草本和灌木组成的下层植物的竞争加剧,从而减弱了幼苗的生长,特别是在土壤营养含量低的立地上(Jokela et al., 2000; McGuire et al., 2001)。
3.4 林隙年龄对更新的影响林隙年龄是林隙动态特征的一个重要变量。随着林隙形成年龄的增加,林隙内的无机环境和生物环境都会有不同程度地变化,不同的树种则产生不同的更新反应,在资源利用和竞争中形成各自的生态位。由表 2可以看出,随着林隙形成时间的延长,各特征指标均有较大增长,其中LAI增加了99.3%,幼苗平均高增加了72.5%,平均基径增加了44.4%,个体密度增加了36.4%,树种数增加了26.1%。可见幼苗的叶面积和苗高生长最快。对比林隙内外的更新状况可以看出,除种密度外,林隙内的各特征指标均优于对照林分,且随着林隙形成时间的延长,这种优势愈加明显。其中个体密度和LAI尤为显著,表明采伐干扰形成的林隙环境更有利于幼苗的繁殖和生长。这是因为采伐林隙形成后,林隙内的光照增加,竞争减少,环境异质性加大,为各类树种的萌发、定居和生长提供了多样化的空间。种子库中的种子大量萌发,幼苗的生长状况也有较大提高。张春雨等(2006)的研究表明,随着时间推移,林隙内土壤水分含量增加,养分循环加快。因此,随着林隙形成时间的增加,林隙内的资源有效性大大提高,进一步促进了幼苗的繁殖和生长。
|
|

林隙内不同位置更新幼苗的特征见图 3。由图 3可见,林隙内不同位置的更新差异明显。3龄林隙内,幼苗种密度、个体密度、平均高和平均基径的最大值均位于林隙西缘(WW),分别为1.25种·m-2、2.80株·m-2、1.11 m和1.0 cm,种密度和个体密度的最小值位于林隙中东部(CE),分别为0.5种·m-2和1.1株·m-2,为最大值的40%和39.3%;平均高的最小值位于林隙中心(CC),为0.88 m,为最大值的79.3%;平均基径的最小值位于中北部(CN),为0.73 cm,为最大值的73%。9龄林隙中各特征值的最大值和最小值的分布无明显规律,林隙北缘(NN)的种密度最大,为1.19种·m-2,中西部(CW)的个体密度最大,为3.94株·m-2,东缘(EE)的种密度和个体密度均最低,分别为0.75种·m-2和1.94株·m-2,是最大值的63%和49.2%;幼苗平均高在中南部(CS)最大,为1.53 m,在中东部(CE)最小,为0.89 m,是最大值的58.2%;幼苗平均基径在林隙东缘(EE)最大,为1.01 cm,在中西部(CW)最小,为0.67 cm,是最大值的66.3%。9龄林隙内各部位幼苗的个体密度均高于3龄林隙,大多数部位幼苗的种密度和平均高大于3龄林隙。幼苗的小个体使得其对微生境的变化特别敏感,在天然更新过程中, 微生境的有利与否对更新幼苗的发生与存活具有决定性的作用。因此,杨桦次生林林隙内不同位置的更新差异可能与林隙内由光照、凋落物等引起的微生境异质性密切相关(Gagnon et al., 2003)。

|
图 3 林隙内不同部位的更新特征(n=16) Figure 3 Trait of seedlings in different sites in gaps(n=16) |
长白山杨桦次生林经营择伐形成的冠林隙面积一般为50~200 m2,多数呈椭圆形、南北方向。林隙内更新幼苗的种类组成较简单,且各种类间的个体数量分布极不均匀,但更新幼苗的种数、个体密度、LAI、平均高生长和径生长均优于非林隙林分。随着采伐林隙形成时间的延长,更新幼苗的种数、个体密度、平均高和平均基径均有显著增加,生长状况相比非林隙林分有明显提高,林隙更新的优势愈加凸出,说明林隙更新是森林更新的一种重要方式。林隙内不同位置的更新状况存在较大差异,3龄林隙的西缘最好,中心及北部最差,9龄林隙内规律不明显。
干扰发生后,林隙内的环境条件发生了不同程度的变化,一般认为从郁闭的林冠层、林隙边缘直到林隙中心存在一个资源梯度,不同树种的不同个体在对林隙资源的利用和竞争中形成了各自生态位的分化,因此也呈相应的梯度分布。Gagnon等(2003)对美国佛罗里达州西北部长叶松(Pinus palustris)林隙的研究发现,较大的幼苗通常位于林隙内初始存活率较低而资源有效性较高的位置(即林隙中心),而较小的幼苗通常位于林隙内初始存活率较高而资源有效性较低的地方(即林隙边缘)。在美国西部针叶林林隙的研究中也发现了类似现象(Gray et al., 1996)。Zhu等(2003;2005)对日本黑松(Pinus thunbergii)海岸林内间伐产生的3种不同大小林隙(林隙直径与周边林木树高的d/H比值分别为0.5, 1.0, 1.5)的更新进行了研究。结果表明,不同大小林隙的幼苗分布均显示出一个相同的趋势:幼苗的高峰分布区不是在林隙中心,而是靠近林隙的北缘、西缘和东缘,这与日本黑松是不耐荫树种相符合,也印证了林隙分配假说,即北方森林中不耐荫树种的幼苗分布在林隙内的北边缘。本研究同上述学者的研究结果不尽一致,可能与林分类型、林隙形状及更新树种等因素有关。以上研究均表明,由于林隙内存在微生境异质性,更新幼苗的分布和生长也是不均匀的。
林隙大小是林隙的重要特征之一,它影响着林隙内的环境因子以及资源的有效性,进而对各物种的繁殖和生长产生不同的作用,是影响幼苗生长的关键性因素(Meer et al., 1999)。小林隙一般主要由周围树木侧生长来填充,而较大的林隙则主要靠更新树木向上生长来填充。Zhu等(2003)发现,大林隙中日本黑松幼苗的平均高和基径都显著高于小林隙,日本黑松幼苗的生长随林隙增大而显著增加。James等(2005)在热带雨林林隙中的更新试验也表明,大林隙中幼苗的生长明显快得多。大林隙中幼苗生长的更好可能是由于:1)光和水的有效性增加;2)凋落物积累的减少或快速分解。另外,随着林隙的增大,来自林隙边缘成熟木对水和养分的竞争压力也下降了,这对林隙内幼苗的生长也是有利的(Madsen et al., 1997)。但林隙存在一个最大值,林隙面积超过该值后对幼苗生长并不能带来进一步的好处。本研究中,小林隙(50 m2)内的更新与非林隙林分无明显差异,大林隙(>200 m2)不利于幼苗的更新,100~150 m2大小的林隙最适合于幼苗更新,不但个体密度大,而且高生长、径生长、叶面积等指标均达到最优状态。而臧润国等(1999)对原始红松(Pinus koraiensis)阔叶林自然干扰形成的林隙内的更新研究表明,20~40 m2的林隙中乔木树种更新的密度最大,认为这么大的林隙既增加了空间和资源的有效性,同时草本植物和灌木也不太繁茂,竞争作用较小,因而促进了树种的大量更新。2个研究结果之所以存在较大差异,可能是2种群落类型不同,林分特征有较大差别所致。Jurij(2002)通过对林隙微生境和林隙内挪威云杉(Picea abies)幼苗更新状况的研究认为,适宜挪威云杉更新的林隙面积应大于500 m2,但在超过1 000 m2的大林隙中,浓密的地面植被会阻碍挪威云杉的更新。刘庆(2004a;2004b)对滇西北亚高山针叶林林隙更新的研究表明,50~100 m2大小的林隙中云杉(Picea likiangensis)、长苞冷杉(Abies georgei)幼苗的高度最大,生长最快,大于100 m2的林隙中个体最小,生长最慢,但都大于非林隙林分。刘金福等(2003)对常绿阔叶林格氏栲(Castanopsis kawakamii)林林隙更新的研究表明,更新树种的多样性在80~160 m2林隙内最大,在>400 m2的林隙内最小。由此可见,在不同区域、不同类型的森林群落中,林隙更新都存在一个最适面积和阈值面积。幼苗更新在最适林隙面积内达到最优,超过阈值面积则急剧下降。这种林隙更新的最适面积和阈值面积因不同地区的不同森林类型而各异。因此,综合本研究的结果,笔者初步认为100~150 m2是长白山杨桦次生林林隙更新的最适面积,在杨桦次生林的经营改造中应注意控制采伐强度,使采伐干扰形成的林隙面积在100~150 m2左右,以更好地促进杨桦次生林的更新和演替。
由于多种因素的制约,本研究只对更新幼苗的基本生长特征做了调查,这只能说明林隙内幼苗的生长现状,而反映不出幼苗的连年生长状态。今后的研究工作中应对林隙内的每棵幼苗进行编号标记,隔数年复查一次,研究随林隙形成时间的延长,幼苗的更新反应和变化,更全面地了解林隙内幼苗的更新规律。同时,对林隙更新中的优势幼苗还应做植物生理生态方面的观测,如光合作用等,从生理生态角度探讨幼苗对林隙干扰的反应。
安树青, 洪必恭, 李朝阳, 等. 1997. 紫金山次生林林窗植被和环境的研究. 应用生态学报, 8(3): 245-249. DOI:10.3321/j.issn:1001-9332.1997.03.005 |
陈迪马, 潘存德, 刘翠玲, 等. 2005. 影响天山云杉天然更新与幼苗存活的微生境变量分析. 新疆农业大学学报, 28(3): 35-39. DOI:10.3969/j.issn.1007-8614.2005.03.009 |
贺金生, 刘峰, 陈伟烈, 等. 1999. 神农架地区米心水青冈林和锐齿槲栎林群落干扰历史及更新策略. 植物学报, 41(8): 887-892. DOI:10.3321/j.issn:1672-9072.1999.08.019 |
郝清玉, 周玉萍, 孙显林. 1998. 采伐作业产生的大林窗对阔叶林天然更新的影响. 吉林林学院学报, 14(2): 69-73. |
胡理乐, 朱教君, 于立忠, 等. 2007. 一种测量林窗面积的改良方法:等角椭圆扇形法. 生态学杂志, 26(3): 455-460. DOI:10.3321/j.issn:1000-4890.2007.03.029 |
梁晓东, 叶万辉. 2001. 林窗研究进展. 热带亚热带植物学报, 9(4): 355-364. DOI:10.3969/j.issn.1005-3395.2001.04.015 |
刘金福, 于玲, 洪伟, 等. 2003. 格氏栲林林窗物种多样性动态规律的研究. 林业科学, 39(6): 159-164. DOI:10.3321/j.issn:1001-7488.2003.06.027 |
刘庆. 2004a. 林窗大小和位置对丽江云杉自然更新幼苗存活和生长的影响. 应用与环境生物学报, 10(3): 281-285. |
刘庆. 2004b. 林窗对长苞冷杉自然更新幼苗存活和生长的影响. 植物生态学报, 28(2): 204-209. |
刘庆, 吴彦. 2002. 滇西北亚高山针叶林林窗大小与更新的初步分析. 应用与环境生物学报, 8(5): 453-459. DOI:10.3321/j.issn:1006-687X.2002.05.002 |
宋新章, 肖文发. 2006. 林隙微生境及更新研究进展. 林业科学, 42(5): 114-119. |
宋新章, 李冬生, 肖文发, 等. 2007. 长白山次生阔叶林采伐林隙更新研究. 林业科学研究, 20(3): 302-306. DOI:10.3321/j.issn:1001-1498.2007.03.002 |
吴刚. 1998. 长白山红松阔叶混交林林冠空隙树种更新动态规律的研究. 应用生态学报, 9(5): 449-452. |
吴宁. 1999. 贡嘎山东坡亚高山针叶林的林窗动态研究. 植物生态学报, 23(3): 228-237. DOI:10.3321/j.issn:1005-264X.1999.03.005 |
熊小刚, 熊高明, 谢宗强. 2002. 神农架地区常绿落叶阔叶混交林树种更新研究. 生态学报, 22(11): 2001-2005. DOI:10.3321/j.issn:1000-0933.2002.11.029 |
于振良, 于贵瑞, 王秋凤, 等. 2001. 长白山阔叶红松林林隙特征及对树种更新的影响. 资源科学, 23(6): 64-68. DOI:10.3321/j.issn:1007-7588.2001.06.014 |
臧润国, 安树青, 陶建平, 等. 2004. 海南岛热带林生物多样性维持机制. 北京: 科学出版社.
|
臧润国, 蒋有绪, 杨彦承. 2001. 海南岛霸王岭热带山地雨林林隙更新生态位的研究. 林业科学研究, 14(1): 17-22. DOI:10.3321/j.issn:1001-1498.2001.01.003 |
臧润国, 蒋有绪, 余世孝. 2002. 海南霸王岭热带山地雨林森林循环与树种多样性动态. 生态学报, 22(1): 24-32. DOI:10.3321/j.issn:1000-0933.2002.01.004 |
臧润国, 刘涛, 郭忠凌, 等. 1998a. 长白山自然保护区阔叶红松林林隙干扰状况的研究. 植物生态学报, 22(2): 135-142. |
臧润国, 徐化成. 1998b. 林隙干扰研究进展. 林业科学, 34(1): 90-98. |
臧润国, 徐化成, 高文韬. 1999. 红松阔叶林主要树种对林隙大小及其发育阶段更新反应规律的研究. 林业科学, 35(3): 90-98. |
张春雨, 赵秀海, 郑景明. 2006. 长白山阔叶红松林林隙与林下土壤性质对比研究. 林业科学研究, 19(3): 347-352. DOI:10.3321/j.issn:1001-1498.2006.03.016 |
朱教君. 2002. 次生林经营基础研究进展. 应用生态学报, 13(12): 1689-1694. DOI:10.3321/j.issn:1001-9332.2002.12.040 |
Gagnon J L, Jokela E J, Moser W K, et al. 2003. Dynamics of artificial regeneration in gaps within a longleaf pine flatwoods ecosystem. Forest Ecology and Management, 172: 133-144. DOI:10.1016/S0378-1127(01)00808-8 |
Gray A N, Spies T A. 1996. Gap size, within-gap position and canopy structure effects on conifer seedling establishment. Ecology, 84: 635-645. DOI:10.2307/2261327 |
James G R, Matthew L, Mark S A, et al. 2005. Growth response by big-leaf mahogany (Swietenia macrophylla) advance seedling regeneration to overhead canopy release in southeast Para, Brazil. Forest Ecology and Management, 204: 399-412. DOI:10.1016/j.foreco.2004.09.013 |
Jennings S B, Brown N D, Shell D. 1999. Assessing forest canopies and understorey illumination: canopy closure, canopy cover and other measures. Forestry, 72: 59-74. DOI:10.1093/forestry/72.1.59 |
Jokela E J, Martin T A. 2000. Effects of ontogeny and soil nutrient supply on production, allocation, and leaf area efficiency in loblolly and slash pine stands. Canadian Journal of Forest Research, 30: 1511-1524. DOI:10.1139/x00-082 |
Jurij D. 2002. Regeneration dynamics in a Norway spruce plantation on a silver fir-beech forest site in the Slovenian Alps. Forest Ecology and Management, 161: 27-38. DOI:10.1016/S0378-1127(01)00492-3 |
Lertzman K P, Sutherland G D, Inselberg A. 1996. Canopy gaps and the landscape mosaic in a coastal temperate rain forest. Ecology, 77: 1254-1270. DOI:10.2307/2265594 |
Madsen P, Larsen J B. 1997. Natural regeneration of beech (Fagus sylvatica L.) with respect to canopy density, soil moisture and soil carbon content. Forest Ecology and Management, 97: 103-113. |
McGuire J P, Mitchell R J, Moser E B, et al. 2001. Gaps in a gappy forest:plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Canadian Journal of Forest Research, 31: 765-778. DOI:10.1139/x01-003 |
Mclaren B E, Janke R A. 1996. Seedbed and canopy cover effects on balsam fir seedling establishment in Isle Royal National Park. Canadian Journal of Forest Research, 26: 782-793. DOI:10.1139/x26-088 |
Meer P V D, Digan P, Savench A G. 1999. Effect of gap size on seedling establishment, growth and survival at 3 years in mountain ash (Eucalyptus regnans F. Muell.) forest in Victoria, Australia. Forest Ecology and Management, 117: 33-42. DOI:10.1016/S0378-1127(98)00471-X |
Myers G P, Newton A C, Melgarejo O. 2000. The influence of canopy gap size on natural regeneration of Brazil nut (Bertholletia excelsa) in Bolvia. Forest Ecology and Management, 127: 119-128. DOI:10.1016/S0378-1127(99)00124-3 |
Runkle J R. 1998. Changes in Southern Appalachian canopy tree gaps sampled thrice. Ecology, 79: 1768-1780. DOI:10.1890/0012-9658(1998)079[1768:CISACT]2.0.CO;2 |
Runkle J R. 1982. Patterns of disturbance in some old-growth mesic forests of eastern north America. Ecology, 63(5): 1533-1546. DOI:10.2307/1938878 |
Sipe T W, Bazzaz F A. 1995. Gap partitioning among maples(Acer) in central new England: survival and growth. Ecology, 76: 1587-1603. DOI:10.2307/1938160 |
Spies T, Franklin J F. 1989. Gap characteristics and vegetation response in coniferous forests of the Pacific northwest. Ecology, 70: 543-545. DOI:10.2307/1940198 |
Zhu J J, Kang H Z, Tan H, et al. 2005. Regeneration characteristics of natural Mongolia pine (Pinus slvestris var. mongolica) forests on sandy land in Honghuaerji. Journal of Forestry Research, 16(4): 253-259. DOI:10.1007/BF02858184 |
Zhu J J, Matsuzaki T, Lee F Q, et al. 2003. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. Forest Ecology and Management, 182: 339-354. DOI:10.1016/S0378-1127(03)00094-X |