 2008, Vol. 44
2008, Vol. 44文章信息
- 赵建兴, 杨忠岐, 任晓红, 梁小明.
- Zhao Jianxing, Yang Zhongqi, Ren Xiaohong, Liang Xiaoming.
- 红脂大小蠹的生物学特性及在我国的发生规律
- Biological Characteristics and Occurring Law of Dendroctonus valens in China
- 林业科学, 2008, 44(2): 99-105.
- Scientia Silvae Sinicae, 2008, 44(2): 99-105.
-
文章历史
- 收稿日期:2007-02-28
-
作者相关文章


2. 内蒙古农业大学农学院 呼和浩特 010019;
3. 山西省安泽县林业局 安泽 030600;
4. 山西省关帝山林管局白虎岭林场 汾阳 043003
2. College of Agronomy, Inner Mongolia Agricultural University Huhhot 010019;
3. Forestry Bureau of Anze County Anze 030600;
4. Baihuling Forest Farm of Guandishan Forest Management Bureau, Shanxi Province Fenyang 043003
红脂大小蠹(Dendroctonus valens,RTB)属鞘翅目小蠹科(Coleoptera:Scolytidae),大小蠹属(Dendroctonus)昆虫,是松杉类针叶树种的蛀干害虫,原产地为美国和加拿大等北美地区,为害松属、云杉属、黄杉属、冷杉属和落叶松属的超过35种植物的根部和干部(Smith, 1971)。自从20世纪90年代末期在我国山西省发现该害虫以来,在短短的几年里,红脂大小蠹为害面积迅速扩大。到2002年,已经在包括山西、河北、河南和陕西四省的52.6万hm2的油松(Pinus tabulaeformis)地严重发生,超过3 500万株油松致死,损失25亿元,已经成为我国森林的第二号杀手(苗振旺等,2002)。2005年又在北京市的门头沟区发现该虫。其已经对我国的油松造成严重为害,成为我国重大的外来入侵害虫之一。
经过专家鉴定,RTB为我国新纪录种,根据其发生历史的追踪调查推测认为,中国的红脂大小蠹可能是20世纪80年代中期随着从北美进口的带皮松材从美国的西海岸传入我国山西省的(李计顺等,2001;Britton et al., 2002)。这个推测已经被基因多样性的研究所证实,即中国的红脂大小蠹基因型与华盛顿俄勒冈州和加利福尼亚州的基因型很相似(Cognate et al., 2005)。经过害虫危险性分析(PRA)方法评价,红脂大小蠹在中国属于高度危险的森林有害生物(宋玉双等,2000)。
红脂大小蠹在我国最早于1998年7月在山西省晋城、沁水地区发现,在当年严重侵害健康油松。随后几年,该虫迅速扩散、蔓延。研究发现,红脂大小蠹以成虫和幼虫在油松树干基部和根部的皮下取食为害,造成大量油松衰弱和死亡。国家林业局2000年开始将此虫列为六大病虫害治理工程。该工程项目实施以来,各地采用药剂熏杀、信息素诱杀成虫等措施进行红脂大小蠹的防治,取得了一定的防治效果。
自红脂大小蠹传入我国以来,有多个作者对其发生规律和生活习性进行了深入的研究和报道(常宝山等,2001;苗振旺等,2001;张历燕等,2002;李同利,2005;王平等,2005;贺虹等,2005),各发生地区报道的红脂大小蠹年发生世代和发生时期不同,各作者报道的红脂大小蠹的生物学习性也有差异。
各地报道的红脂大小蠹的生活习性和发生规律有很多的不同之处,同时对红脂大小蠹的某些特性还缺乏清楚、准确的了解,这对于害虫的防治和天敌的使用非常不利。为了进一步明确红脂大小蠹的生活规律和习性,笔者在山西、河北省的几个地区做了连续的详细观察,对红脂大小蠹的某些行为习性和发生规律进行了深入的观察和模拟试验,以验证、补充对这种入侵害虫的了解。
1 材料与方法 1.1 红脂大小蠹中国种群的生物学特性观察 1.1.1 越冬场所、部位和虫态调查2003年11月、2004年3月上旬和5月,分别在灵空山林场、白虎岭林场、芦芽山自然保护区油松林的受害树和伐桩的干部和根部、伐木的树皮下检查红脂大小蠹的越冬部位、越冬虫态和存活情况。
1.1.2 野外寄主调查和室内胁迫饲喂试验2003年在灵空山林场,对采自林间的红脂大小蠹成虫和幼虫,分别饲喂当地不同树种的形成层树皮,观察其取食和产卵情况。
在分布其他松树和杉树的红脂大小蠹发生地区,各年度进行踏查,以了解野外红脂大小蠹的寄主情况。
1.1.3 为害坑道和生长发育观察在室内选取40 cm树段,两端封蜡,放入整理箱中,分别接入2~3对红脂大小蠹成虫,分别在第20、40、60天打开部分受害坑道,观察坑道情况和害虫生长发育情况。
在各地林间,在不同时期分别解剖寄主受害坑道,观察坑道结构、种群结构、为害状态和存活情况。
1.1.4 其他习性观察结合夹板饲养、树段饲养和林间观察等手段,对红脂大小蠹的其他生活习性和与其他昆虫的关系进行观察。
1.2 红脂大小蠹在中国发生区发生规律的研究 1.2.1 年生活史的系统观查2003年从红脂大小蠹发生盛期的8月至11月中旬,在太岳山林管局灵空山林场野峪口和孔房峪林地,标记100株当年新受害油松,每3天选取2~3个红脂大小蠹坑道,观察其中的种群结构和数量,树体症状等。另外结合各林场红脂大小蠹的诱捕器监测情况,各地寄主受害率和新侵入孔情况,分析各年份和各地区的发生时间和发生程度。
2005年在山西省晋中市榆次区庆城林场,网罩红脂大小蠹为害坑道,系统观察害虫在林间的发育情况。另外,在红脂大小蠹的越冬期前后,在山西南部、中部、北部选择代表性林地进行越冬场所和越冬虫态调查。
1.2.2 发生程度和环境因素调查从2003年到2005年,通过不定点踏查的方式,在红脂大小蠹越冬期和发生盛期,分别在河北省井陉县、邢台市内丘县、邯郸市武安县,河南省的修武县、辉县,陕西省的黄龙地区大岭林场、韩城市雷寺庄林场,山西省的安泽县良马林场、太岳山林管局灵空山林场、太行山林管局禅堂寺林场、关帝山林局的白虎岭林场和三道川林场、管涔林局的接官亭林场和芦芽山自然保护区调查油松受害程度、其他树种是否受害、各时期害虫的主要发生虫态、各林地特征等情况,分析红脂大小蠹发生的主要影响因素。
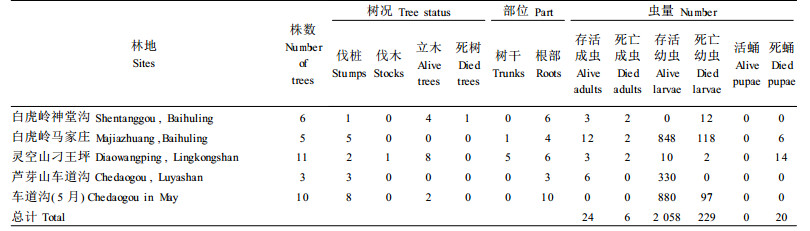
2 结果与分析 2.1 红脂大小蠹中国种群的生物学特性 2.1.1 红脂大小蠹的越冬部位和虫态调查发现,各地红脂大小蠹的越冬场所和虫态有很大的不同,结果见表 1。从表 1可知,在白虎岭、灵空山和芦芽山等林地,红脂大小蠹主要的越冬场所是伐桩,占调查发现有虫株的54.3%,其次是活立木、死树和偷伐木;以大龄幼虫和少量的成虫和蛹越冬,安全越冬部位主要在伐桩的根部。在白虎岭林场,越冬虫态为老熟幼虫和少量蛹、成虫,主要在伐桩主根和侧根皮层下,多数已经做好蛹室;芦芽山地区越冬虫态主要为2~3龄幼虫,也有少数老熟幼虫和部分蛹、成虫,群集在主根部位;灵空山则主要以2~3龄幼虫在寄主根部,少量成虫和幼虫可以在伐木下越冬,部分幼虫和成虫在干部坑道中越冬,但是不能存活。在各地区的不同林地,红脂大小蠹的越冬虫量也有较大差异,如白虎岭林场的神堂沟和马家庄伐桩根下的虫量明显不同,这也说明红脂大小蠹越冬时,当代成虫产卵量很不一致。
|
|
根据红脂大小蠹在北美地区对松属、杉属、落叶松属等多种树种的为害情况,笔者在室内对在我国红脂大小蠹发生区生长的几种主要树种做了树皮饲喂试验,结果见表 2。
|
|

从表 2可以看到,在胁迫条件下,红脂大小蠹成虫可以取食油松和华山松韧皮部,对华北落叶松和云杉有少量的取食,对圆柏和山杨则完全不取食;幼虫则除了油松外,只少量取食了华山松,对圆柏和山杨则完全不取食。但是成虫和幼虫在不取食的情况下,仍然可以存活半个月左右。这表明,红脂大小蠹一旦被带到无适宜寄主的地区仍然可以存活相当的时间,提示生产中对红脂大小蠹的检疫是十分必要的。
在野外红脂大小蠹的寄主调查中,2003年在太岳山林管局的灵空山林场全年的其他寄主调查中,没有发现华山松和白皮松受害。在2004年和2005年陕西省韩城市雷寺庄的白皮松林地调查中,也没有发现白皮松受害(但是当地技术人员反映,在前几年偶有发现);2005年在榆次区乌金山林场,发现部分白皮松有侵害,症状如油松,但是漏斗状侵入孔不明显;其他树种未发现被红脂大小蠹侵害。在各地的华北落叶松林地调查,都没有发现红脂大小蠹的为害。
2.1.3 红脂大小蠹坑道的建立和生长发育2004年和2005年分别在布鲁塞尔自由大学实验室和国内的庆城林场室内,用黑松和油松树段,在22 ℃左右温度饲养情况下,配对的红脂大小蠹成虫在当天就可以挖掘坑道,通常先横向挖掘2~3 cm的一段,然后向下建立母坑道。2~30天后产卵,卵通常产在坑道一侧,然后用木屑和虫粪覆盖卵块,成虫继续挖掘坑道,也可以再次产卵。试验显示,在接入成虫35天左右,红脂大小蠹幼虫已达2龄;60天左右,部分幼虫已经老熟、做蛹室;90天后,新羽化成虫占61%,幼虫占20.5%, 蛹18.5%,表明绝大部分昆虫在适宜温度下,3个月内可以完成1代。
林间观察,在山西省晋中市榆次庆城林场的野外笼罩试验中,从2005年7月15日—8月22日,14个有红脂大小蠹成虫侵入孔的为害坑道中,有7个坑道被红脂大小蠹成功定居,占50%,平均每个坑道幼虫87.3头,55%为低龄幼虫,43%为卵期,2%为成虫(越冬代成虫)。比较上述室内的发育进度可以看出,由于林间是变温条件,在林间红脂大小蠹的发育要比室内慢得多。
在树段饲养中可以看到,在成虫成功侵入寄主并且产生后代后,部分成虫有钻出饲养树段重新钻蛀的现象,表明当寄主已经被害虫成功繁育后代后,成虫可以转主为害(图 1、图 2)。

|
图 1 树段粗皮下的红脂大小蠹排粪孔 Figure 1 Hole of RTB on the stock of P. tabulaeformis |

|
图 2 红脂大小蠹排粪孔的形状和距离 Figure 2 Shape and distance of holes |
成虫堵塞坑道入口:试验室内和林间观察发现,在红脂大小蠹进入坑道后,它们不断地将碎屑、松脂和虫粪从侵入孔排出,在侵入孔外形成柔软的凝脂。当它们交尾、产卵后,用虫粪及碎屑覆盖卵块,然后依然继续从侵入孔排出碎屑和虫粪,直至坑道过深才用虫粪堵塞坑道口,在坑道入口下的10~14 cm处以下从内部咬出2~5个数目不等的新排粪孔,但是并不咬通,留下外部粗皮。当大量幼虫产生的虫粪和成虫虫粪太多时,才咬通排粪孔,由成虫将虫粪推出坑道。成虫伴随幼虫生活,或者钻出坑道(在室内树段饲养中多见),或者死在坑道中。在树段饲养中,老成虫经常可以活到新成虫羽化以后,说明很多成虫寿命可以在100天以上(图 3、图 4)。

|
图 3 RTB成虫坑道中的卵、虫粪、母坑道位置 Figure 3 RTB eggs and frass in gallery |

|
图 4 RTB幼虫孵化后与成虫共同生活的坑道 Figure 4 RTB larvae and adults living together in gallery |
通过3年多的系统观察和不定期的踏查,笔者观察到,在山西省太岳山林区以北的红脂大小蠹发生地区,河北的所有发生区,河南的多数发生区都为1年发生1代,从5月底到9月林间的主要虫态为成虫、卵或低龄幼虫;山西的安泽以南地区,陕西大岭、韩城地区和河南辉县可以存在部分1年发生2代的种群,这些都是主要以成虫越冬的种群。无论哪个地区,在主要生长季节林间一直存在成虫的扩散,表明以幼虫越冬的种群发育不整齐,造成除了初夏的扬飞高峰外,林间以1代发生为主的现象。
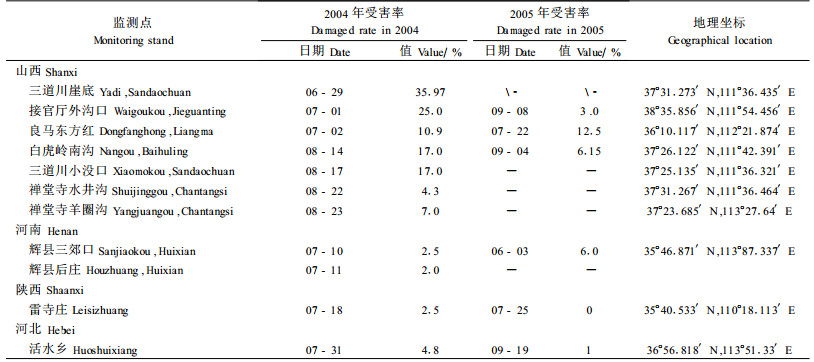
2.2.2 发生程度和环境因素结合各地固定观测点和各监测点不同时期林间的寄主受害率、平均受害坑道数量和林地状况等调查情况,结合几年来各地的气象资料和地理环境差异情况,笔者分析了红脂大小蠹发生与寄主状况、林分、地理位置的关系,如表 3所示。
|
|
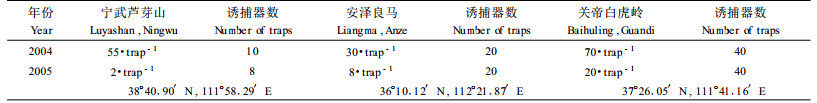
从3个林地诱捕器诱捕到的红脂大小蠹成虫的数量比较可以看到,各地2005年的害虫种群数量下降明显(表 3)。但是各地的下降幅度有很大的差别,芦芽山的下降幅度最大,其次是白虎岭林场,良马林场的下降幅度较小,其主要原因是,2005年5月在该林地发生了火灾,烧毁了大片油松,并且在随后进行了砍伐。林地残留大量的伐桩和过火木,这有利于红脂大小蠹安全度过早春的寒冷气候和吸引越冬后成虫的侵害,这与笔者后期在林间的调查是吻合的(表 3,4,图 5)。其他林地的下降主要是由于气候因素和各地采取了更好的保护措施,林地中新伐桩减少不利于红脂大小蠹的大量繁殖和安全越冬。
|
|

|
图 5 固定观察点油松受害率 Figure 5 Ratios of RTB attacked trees in different monitoring sites |
2005年红脂大小蠹发生季节,笔者分别赴山西、河南、陕西、河北的部分害虫监测点调查,红脂大小蠹发生情况见表 4。
由于时间、交通工具、其他任务等多种原因笔者没有能对大部分的观测点进行连续的详细调查,有待以后进行连续年份的监测。
3 结论与讨论经过历时4年多的林间调查和试验室饲养观察,笔者对红脂大小蠹在中国的主要发生区的主要生物学习性有了比较清晰的了解。
1) 在我国各主要发生地,红脂大小蠹都是主要在伐桩、衰弱树和新死亡树的根部以大龄或老熟幼虫和少量成虫、蛹越冬为主,部分地区也可以以成虫、老熟幼虫在伐木皮下越冬。来年越冬虫态羽化为成虫后,陆续从树基部旧坑道的羽化孔或直接从地下钻出,扬飞扩散。这为采取更加经济有效的防治措施提供了依据。
本研究的一个重要发现是:在山西省南部太岳山林区以南、河北和河南、陕西南部的部分发生区,伐木树皮下也是红脂大小蠹的重要越冬场所之一,这对了解当年害虫发生来源、压低初期种群数量具有重要的意义。因此对这些地区没收的盗伐受害木和自用木材要在堆积场所进行剥皮或者药剂处理,避免皮下的害虫越冬后成为来年的侵害来源。
2) 红脂大小蠹在我国山西省、河南省、河北省、陕西省发生区不同地区可以1年发生1~2代,以成虫、幼虫或蛹在树干基部、主根、侧根皮下越冬,世代重叠。这与多位作者前期研究的结果相吻合。张历燕等(2002)观察,该虫在山西沁源、太岳山林区海拔1 100~1 700 m的地区以成虫(占25%)、老熟幼虫(占52.5%)、3龄幼虫(占18.2%)、2龄幼虫(占3.5%)以及少量蛹(占0.8%)在树干基部、主根、侧根的韧皮部越冬。这与笔者在各地林地调查的情况是吻合的。60%~70%的为1年1代,30%~40%为3年2代,或2年1代。随着海拔的升高,以2~3龄越冬的个体数逐渐减少。除冬季见不到卵外,其他虫态在全年均可见到。
常宝山等(2001)观察,该虫在晋城地区1年发生2代,以成虫、蛹、幼虫在树干基部和主、侧根皮层的取食坑道内越冬。成虫3月中旬开始活动,4月下旬开始侵入寄主,5月上中旬为越冬代成虫扬飞、侵入和产卵盛期,8月中下旬为第1代成虫扬飞、侵入和产卵盛期,10月下旬第2代成虫开始羽化。在温度、湿度和寄主等因子的共同影响下,其发育不太整齐,有世代重叠现象。
在河南、河北、陕西的红脂大小蠹发生区,分别报道1年发生1代或者2代,或者2年1代(李同利,2005;王平等,2005;贺虹等,2005),多数地区以成虫和大龄幼虫在油松根部越冬。
3) 对红脂大小蠹寄主的调查和试验,虽然还没有发现在中国发生区已经报道的树种以外的寄主受害,但是,它们很强的耐饥饿能力和高扩散能力仍然提示人们,在调运种苗、带皮木材时,要做好严格的检疫措施,避免人为的红脂大小蠹助迁。
4) 在害虫发生区受害程度的调查比较中可以明显地看到,火灾、过度割松脂作业和乱砍乱伐等人为不良因素存在的林地,红脂大小蠹发生程度重于其他林地,特别在2005年各地红脂大小蠹发生普遍大幅下降的情况下,安泽县良马林场花豹沟林地的发生却重于2004年,当年5月的严重火灾是这种现象的直接原因。因此,做好林地的防火、防盗伐是控制红脂大小蠹发生非常有效的措施。Yan等(2005)认为,20世纪80年代以来,由于中国从美国大量连续进口西松原木,随木材带来的红脂大小蠹在新的生境下重复累积,并且红脂大小蠹的美国和中国种群的DNA序列研究说明,从美国多来源的种群提供了在新生境成功定居和入侵的可能性,造成了入侵种群更多的遗传变异。另外,红脂大小蠹的高繁殖率、短的生活史、表型可塑性和高的扩散能力等特点使之更容易适应不同的环境压力,这些可以解释该虫在中国部分地区的成功定殖。另外,在中国由于红脂大小蠹种群缺乏种间竞争,存在丰富的食物资源、有效天敌的缺乏都造成了害虫暴发的基础(Yan et al., 2005)。
红脂大小蠹成虫侵入坑道的一系列习性为采取防治措施和运用天敌控制红脂大小蠹的可能提供了有力的支持,熏蒸杀虫剂和天敌进入害虫坑道的途径是始终存在的。
还有一些红脂大小蠹的习性还不是很清楚,比如在自然情况下红脂大小蠹成虫是否存在转主为害,如果转主是否重新寻找配对等还有待于深入研究。
常宝山, 刘随存, 赵小梅, 等. 2001. 红脂大小蠹发生规律研究. 山西林业科技, (4): 1-4. DOI:10.3969/j.issn.1007-726X.2001.04.001 |
高宝嘉, 信金娜, 关慧元, 等. 2003. 红脂大小蠹的发生和为害规律. 动物学杂志, 38(5): 71-73. DOI:10.3969/j.issn.0250-3263.2003.05.016 |
郭泉水, 徐德应, 阎洪. 1995. 气候变化对油松地理分布影响的研究. 林业科技, (5): 394-401. |
贺虹, 李梦楼, 郭新容, 等. 2005. 红脂大小蠹生物学特性研究. 西北林学院学报, 20(1): 140-142. DOI:10.3969/j.issn.1001-7461.2005.01.035 |
李同利. 2005. 红脂大小蠹的特征及综合防治. 河北林业科技, (6): 46. DOI:10.3969/j.issn.1002-3356.2005.06.025 |
李计顺, 常国彬, 宋玉双, 等. 2001. 实施工程治理控制红脂大小蠹虫灾——对红脂大小蠹暴发成因及治理对策的探讨. 中国森林病虫, (4): 41-44. DOI:10.3969/j.issn.1671-0886.2001.04.018 |
李鹏飞, 张跃宁, 孙永康, 等. 2005. 红脂大小蠹的种群动态及预测预报. 西北林学院学报, 20(3): 116-119. DOI:10.3969/j.issn.1001-7461.2005.03.030 |
苗振旺, 周维民, 霍履远, 等. 2001. 强大小蠹生物学特性研究. 山西林业科技, (4): 1-4. DOI:10.3969/j.issn.1007-726X.2001.04.001 |
苗振旺, 赵明梅, 卢学林. 2002. 大小蠹植物引诱剂对红脂大小蠹诱引效果试验. 山东林业科技, (1): 23-25. DOI:10.3969/j.issn.1002-2724.2002.01.010 |
宋玉双, 杨安龙, 何嫩江. 2000. 森林有害生物红脂大小蠹的危险性分析. 森林病虫通讯, (6): 34-37. DOI:10.3969/j.issn.1671-0886.2000.06.014 |
王平, 李书吉, 姚随印, 等. 2005. 红脂大小蠹生活史及生物学特性观察. 河南林业科技, 25(2): 15-17. |
王鸿斌, 张真, 孔祥波. 2005. 油松萜烯类挥发物释放规律与红脂大小蠹危害的关系. 北京林业大学学报, 27(2): 75-80. DOI:10.3321/j.issn:1000-1522.2005.02.015 |
许佳林, 王建军, 李红平, 等. 2002. 红脂大小蠹越冬场所及成虫出土观察. 山西林业科技, (4): 26-28. DOI:10.3969/j.issn.1007-726X.2002.04.006 |
张历燕, 陈庆昌, 张小波. 2002. 红脂大小蠹形态学特征及生物学特性研究. 林业科学, 38(4): 95-99. DOI:10.3321/j.issn:1001-7488.2002.04.016 |
Britton K O, Sun J H. 2002. Unwelcome guests: Exotic forest pests. Acta Entomologica Sinica, 45(1): 121-130. |
Cognato A I, Sun J H, Anducho-Reyes M A, et al. 2005. Genetic variation and origin of red turpentine beetle (Dendroctonus valens LeConte) introduced to the People's Republic of China. Agricultural and Forest Entomology, 7: 87-94. DOI:10.1111/afe.2005.7.issue-1 |
Smith R H. 1971. Red turpentine Beetle. USDA Forest Service Forest Pest. Leaflet, 55: 8. |
Sun J H, Miao Z W, Zhang Z N, et al. 2004. Red turpentine beetle, Dendroctonus valens LeConte (Coleoptera:Scolytidae), response to host semiochemicals in China. Environmental Entomology, 33(2): 206-212. DOI:10.1603/0046-225X-33.2.206 |
Yan Zhengliang, Sun Jianghua, Donnand R O, et al. 2005. The red turpentine beetle, Dendroctonus valens LeConte (Scolytidae):an exotic invasive pest of pine in China. Biodiversity and Conservation, 14: 1735-1760. DOI:10.1007/s10531-004-0697-9 |