 2008, Vol. 44
2008, Vol. 44文章信息
- 刘仁林, 王琅, 杜天真.
- Liu Renlin, Wang Lang, Du Tianzhen.
- 井冈山罗浮栲群落种群生态数量场特征
- Features of Population Ecological Quantity Field of Castanopsis fabri Community in Jinggang Mountain
- 林业科学, 2008, 44(2): 1-7.
- Scientia Silvae Sinicae, 2008, 44(2): 1-7.
-
文章历史
- 收稿日期:2007-06-07
-
作者相关文章



2. 江西庐山自然保护区管理处 庐山 332900;
3. 江西农业大学园林与艺术学院 南昌 330045
2. The Office of Lushan Mountain Natural Reserve Lushan 332900;
3. College of Landscape Architecture and Art, Jiangxi Agricultural University Nanchang 330045
罗浮栲(Castanopsis fabri)是亚热带常绿阔叶林的优势树种和建群种之一,在我国主要分布于四川、云南及长江以南亚热带海拔350~900 m地区,在森林生态系统中具有重要地位。井冈山位于江西省西南部,森林植被为亚热带常绿阔叶林,其中罗浮栲林是最为常见的森林类型之一,主要分布在沟谷和山坡中、下部。以前主要侧重于栲类群落的区系、外貌及一般群落结构的研究,如邓贤兰(2003)和邓贤兰等(2003;2005)研究了井冈山自然保护区的栲树群落、井冈山自然保护区栲属群落优势种群的种间联结关系和井冈山自然保护区栲属群落植物区系;林英等(1990)研究了井冈山罗浮栲林的群落结构。Gates(1980)出版了第1部物理生态学专著, 使物理生态学成为生态学的一个重要研究领域,许多学者从不同角度探讨生态场理论与方法,如:Ronald等(1985)用生态场理论、方法研究了珊瑚礁鱼类的分布、丰富度和繁殖连续体;Sharpe等(1986)对树木生长的空间补偿进行了物理生态学模拟研究;Penridge等(1988)对树冠光生态场的分布进行了研究;Walker等(1988)探讨了生态场的概念与方法;Masaki等(1999)进行了光、温生态场因子对褐藻(Ecklonia cava)繁殖的分析;Wildova′等(2007)研究了变换生态场参数模拟植物无性系结构与生长的差异等。我国对植物生态场的研究主要有:邹春静等(1999)研究了生态场理论在沙地云杉(Picea meyeri var. mongolica)种群更新中的应用;董振堂等(1989)研究了红松(Pinus koraiensis)种群生态场特性函数;丛沛桐等(2001)研究了东灵山辽东栎(Quercus liaotungensis)林植物生态场;王亚秋等(2005)研究了改进的植物生态场模型与实例分析;蔺银鼎等(2006)研究了城市绿地空间结构对绿地生态场的影响等。
生态场是生物的生命过程与环境相互作用产生的综合生态效应的空间分布(王德利,1994)。在这个定义中,“生命过程”是分不同等级的,如不同的细胞、器官、物种等;“综合生态效应”实质上是含有交互作用的各种生态因子的瞬时状态,它是时间的函数。由于交互作用的复杂性,所产生的“生态效应”即为“综合”的“生态效应”。交互作用是不同生命形式或等级互相依存、相互协同的非线形作用,因此2个类型的群落在一起所产生的“综合生态效应”不等于各自独立所产生的“综合生态效应”的简单叠加。以前考察一个独立的群落,主要注意它对周围其他生命形式和环境的作用,类似于把一个带电体看成对周围的影响,早期的生态场研究都与此类似。随着科学技术的发展和对生态场认识的深入,现在认为,考察森林生态系统中一个群落的动态变化,可以把这片群落看作是生态场中的一个“质点”,这个质点受到整个系统所形成的生态场的强烈作用,而质点本身对生态场产生的作用不是主导地位(Law et al., 2003),类似于把一个星体放在宇宙的引力场中来考察它的运动状态。群落优势种群的数量场就是把群落放在生态系统(包含群落本身)所形成的生态场中,由于群落受到生态场的作用和自身作用,因此种群数量特征的空间分布就形成了数量场,本文基于这种观点,试从生态场的角度研究罗浮栲林优势种群的数量动态规律,为开展罗浮栲林生态系统研究,合理和持续利用资源提供科学依据。
1 试验地概况试验地位于井冈山自然保护区26°32′22″ N,114°08′31″ E,属罗霄山脉中部,山坡下部,群落面积1 200 m2;海拔660 m,坡度为28°。属中亚热带季风型气候,年均气温14.3 ℃,极端最低气温-11 ℃,最高气温40 ℃;年均降水量1 855 mm,4—8月降水量约占全年的63%;年均相对湿度85%。土壤类型主要为山地红壤土,土层较厚,腐殖质丰富。水肥条件均较好,植物种类丰富,林冠呈波浪状,郁闭度0.85。罗浮栲群落的上层以罗浮栲为优势种;除此之外,还有少量的鹿角栲(Castanopsis lamontii)与杉木(Cunninghamia lanceolata)。
2 研究方法 2.1 野外调查用罗盘仪对群落内所有的罗浮栲进行定位测量,测定每一株罗浮栲的方位和到测点的水平距离;除幼苗外,测量相应的胸径、树高和树龄。
2.2 数据处理根据长期的物候观察,罗浮栲大约在树龄25年时开始开花结实(火树华等,1980),因此按25年为1个世代,把罗浮栲群落划分为7个世代,各世代罗浮栲群落数量因子见表 1。
|
|
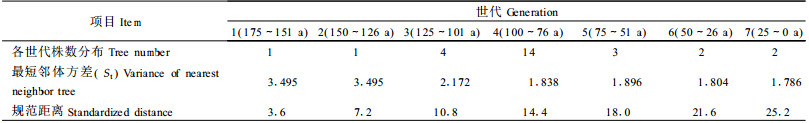
根据每株树的方位和距离,用1/300比例尺绘成空间分布图(图 1),图中括号外的数据为胸径,括号内数据为年龄。根据图 1量出各世代每株树的最短邻体距离,计算各世代的最短邻体距离的方差St(表 1)。由于第1世代和第2世代各只有1株树,因此用群落中最大年龄的3株树的最短邻体距离计算方差,作为这2个世代的St。

|
图 1 罗浮栲种群个体空间分布 Figure 1 Distribution of trees of C. fabri |
以年龄最大的1株罗浮栲作为中心树A,由图 1可知,中心树A到最远1株罗浮栲的距离为24.6 m(苗,3年生),以7个世代等分这段距离(24.6 m),则每个世代的空间等差距离为3.5 m,这样可以依次得到第1至第7个圆的半径分别为3.6、7.2、10.8、14.4、18.0、21.6和25.2 m。为了研究方便,把7个世代依次得到的圆半径(3.6, 7.2, ……, 25.2 m)称为规范距离(谢树艺, 1985)。罗浮栲种群的规范距离的分布见表 1。规范距离反映了一定的生物学特性,以7个世代等分中心树A到最远1株罗浮栲的距离(24.6 m)得到的各世代空间等差距离为3.5 m,这与实测得到的中心树的冠幅半径为3.6 m接近。这个冠幅为罗浮栲群落中最大的冠幅,能满足各世代的个体有充分的生长发育空间。
3 结果与分析 3.1 生态数量场场势函数 3.1.1 生态数量场场势是生态数量场在空间上的变化利用表 1中的St和r能较好地刻画生态数量场的特征,St既反映了罗浮栲种群受到生态场的影响,又反映了种群本身在拓展过程中克服生态场“阻力”潜能。因为种群一般在前面1—2个世代个体的空间分布是随机的,因此St较大;随着种群“拓展”的成功,个体的空间分布由“随机”演化为“团块状”,进而形成大“团块状”,St逐渐缩小,并稳定于某个值;到后期,由于生态场随时间的变化以及种群本身在拓展过程中所造就的环境可能不再适应它自身的生长,从而导致种群数量场场势衰减,规范距离r不再增加。因此罗浮栲群落生态数量场的场势函数可以表示为
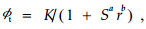
|
(1) |
式中:ϕt为场势函数;K为群落环境支持最大限量;S表示最短邻体距离的方差St;r为规范距离;a、b分别为参数。
用表 1中的St和r对(1)式进行回归拟合,得:
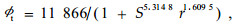
|
(2) |
复相关系数为0.76。用(2)式与Logistic方程Nt=K2(1+e-Rt)比较(式中R为内秉生长率),由于Nt与ϕt都是时间t的函数,所以可写成:

|
(3) |
为了避免左式R与右式中的r混淆,又可把(3)式写成:
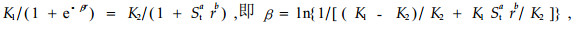
|
(4) |
式中:β为生态数量场内势;K1、K2均为群落环境支持最大限量,且K1=K2。
β反映了罗浮栲种群在生态场中“克服阻力”的能力,即拓展自己的潜力大小,β值愈大,说明种群“克服阻力”的潜能越大(Fabrig, 1988)。在(4)式中,由于K1/K2=1, 根据表 1,按(4)式计算得到表 2。
|
|
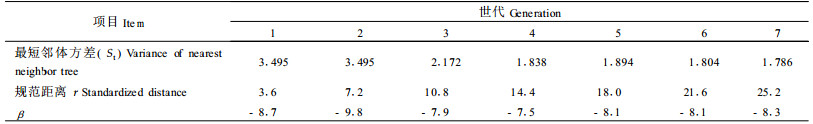
从表 2可以看出,罗浮栲种群第1、2、7世代的β值小于3、4、5和6世代的β值,即“驼峰效应”,说明在群落发展的初期和末期,生态场对罗浮栲种群不利,这2个时期罗浮栲种群克服系统生态场“阻力”的潜能较小,而中间阶段有利于罗浮栲种群的发展。β与物种的遗传基础有关,不同物种β的差异表现为驼峰值的时间不同,如表 2中罗浮栲β驼峰值的时间约100年,大果马蹄荷(Exbucklandia tonkinensis)约75年(刘仁林等,2000),阳性树种可能更低些。
3.1.2 罗浮栲种群生态数量场的作用范围生态数量场作用范围的含义为数量场的边界或有效范围,本质上是种群最大的扩散空间范围。由表 2可知,罗浮栲种群的St大约稳定在1.736左右,因此当r=60 m时,场势ϕt≈0,即数量场的作用范围为60 m(半径),种群最大占有面积可以达到11 304 m2。
3.2 生态数量场梯度罗浮栲种群生态数量场的梯度是数量场场势在生态场中的空间变化及其方向,用公式表示为G=Δϕt/ΔL(式中Δϕt=Δϕt2-Δϕt1,时间t2>t1;ΔL为相邻2个世代的平均规范距离之差)。梯度的特点是具有方向性。由(2)式计算得到1—7世代的ϕt理论值依次为1, 1, 5, 7, 4, 4与3。根据G=Δϕt/ΔL,得到罗浮栲种群1—7世代的生态数量场的梯度在规范距离r(0~25.2 m)上的变化,即规范距离为3.6、7.2、10.8、14.4、18.0、21.6和25.2 m时,梯度G分别为0、1.1、0.6、-0.8、0、-2.8与-2.8。在计算G的过程中,计算第7世代的G值需要知道第8世代的ϕt值(t1=7, t2=8),由(2)式可得到第8世代ϕt的理论值等于2(ΔL=3.6,St=1.736, r=28.8 m)。
梯度为负值表示场势ϕt减弱,正值表示场势ϕt增强。由规范距离与场梯度的相应变化规律可知,罗浮栲种群生态数量场的梯度在规范距离(0~25.2 m)内表现为在3.6~14.4 m场势较强;在18.0~25.2 m场势减弱。实际上图 1也说明了这一点,即以离中心树A的距离0~14.4 m为半径的圆内罗浮栲的个体数量最多,表明这个空间是种群在系统生态场中拓展出的最适合自己生长、发育的生态场。
3.3 生态数量场周期性震荡规律生态场不同于物理场,当把某个生命层次作为生态场研究对象时,必须考虑这个对象所处的生态系统生态场对它的影响和这个对象本身对生态系统产生的影响,这2种关系的相互作用是非线性的、间接的,并且是时间的函数,因此生态场出现周期性震荡现象是客观规律的反映。生态数量场同样具有周期震荡的特征,罗浮栲种群生态数量场也不例外。
3.3.1 罗浮栲种群生态数量场的周期性震荡函数根据表 1各世代的株数得到累进株数列于表 3;此外,根据图 1量出各世代株数中离中心树A最远的一株树的距离,作为这个世代离中心树的最大距离,然后计算各世代累进最大距离(表 3)。
|
|
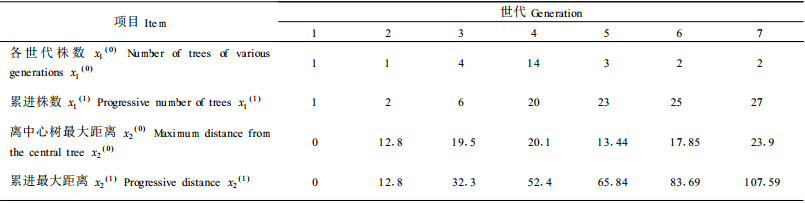
根据表 3,令:
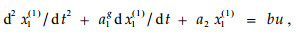
|
参数向量:
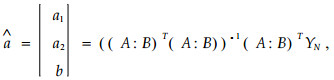
|

|
所以:

|
所以dx1(1)/dt2-0.693 3dx1(1)/dt+1.036x1(1)=0.221u。
系统特征根方程:
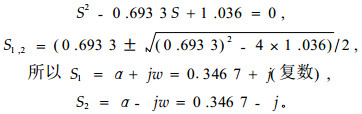
|
由于系统特征根是共轨复数,所以响应函数表示为
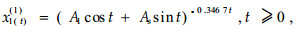
|
式中:x1(t)(1)为系统响应函数;A1、A2为振幅;t为时间(邓聚龙,1985)。
令:
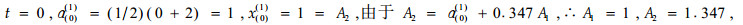
|
故得到
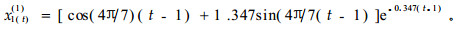
|
(5) |
显而易见,(5)式是一个震荡的衰减过程。分别用t(t=1~16)表示世代作横坐标;用y表示系统响应函数(x1(t)(1))作纵坐标,由(5)式得到周期震荡图 2。

|
图 2 罗浮栲种群数量的震荡曲线 Figure 2 Oscillation of C. fabri population quantity |
由图 2可知,在罗浮栲群落中,罗浮栲优势种群生态数量场是一个周期性震荡的生态场,震荡的周期为2个世代,时间长度为50年。罗浮栲种群生态数量场的震荡与生态场综合因子效应如光、热、水等有关,在正常的森林生态系统中,群落内林窗动态变化可能是这些综合因子效应产生的背景基础,刘金福等(2003)对格氏栲(Castanopsis kawakamii)群落的林窗干扰规律研究时发现,大多数林窗是在50年以前形成的,这与罗浮栲种群数量场的震荡周期基本吻合。
其次,图 2表明,0~10世代振幅较大,时间长度为250年。振幅较大说明罗浮栲种群数量场仍处于“活跃”期,种群数量还有增加。根据调查,罗浮栲的最大年龄为173 a,距250 a约3个世代,因此现在的罗浮栲群落并未进入衰退阶段,这一点从群落的罗浮栲年龄结构(图 1)可以得到检验,群落中年龄结构合理,而且还有幼苗的出现。
此外,从图 2也可知,第13(325年)世代罗浮栲种群数量场衰减很快,并逐渐趋近于0,由此推测罗浮栲群落325年后将进入衰退期,现在的群落优势树种的地位将让位于其他树种。福建三明小湖地区分布有近700 hm2的格氏栲林,约300多年历史,已是成熟的群落,正处于树种更替时期(刘金福等, 2003), 这与罗浮栲群落的周期性基本相似。但是,罗浮栲是常见的亚热带常绿阔叶林优势树种和建群种,前期较耐荫,生长速度较慢,因此罗浮栲群落的衰退期出现得较迟。
图 2所反映的罗浮栲优势种群数量场周期震荡规律,对自然森林群落的动态管理和保护具有重要意义(Maestre et al., 2005),如果需要保护优势种群,可以在震荡周期内辅以人工保育措施和人工促进天然更新措施,延缓衰退期的出现;此外,生态场数量场的周期性震荡规律在人工群落建造和植被恢复中具有实用价值。
4 结论与讨论1) 罗浮栲群落优势种群生态数量场的研究分析表明,植物生态场理论能够科学、准确地描述群落中优势种群的动态规律,揭示种群生态的基本规律,如数量场内势β表达了种群克服生态场“阻力”的潜能和动态规律,这种规律在本质上与种群遗传基础有密切的联系,说明了种群数量场与物种遗传、进化的直接联系;其次,生态数量场梯度G刻画了场势ϕt的强弱变化,揭示了种群如何在生态场中拓展出最适合自己生长、发育的生态空间;此外,系统响应函数x1(t)(1)是种群生态数量场具有周期性震荡的函数,它揭示了生态场中综合生态因子如光、热、水等交互作的规律,综合生态因子的不均质性分布是数量场表现出周期性震荡的主要原因。总之,生态数量场的内势、生态数量场梯度、系统响应函数以及规范距离是揭示种群生态数量场规律的重要因子。
2) 本研究表明,群落生态场的研究具有较严密的数理逻辑基础,表达简洁、准确、实用,能够有效地反映种群动态变化规律的根本原因,是探索群落动态变化的新技术。应用群落生态场的研究方法,可以得到与传统群落动态分析方法如林窗分析模型等基本相同的结果,如刘金福等(2003)研究格氏栲群落的林窗干扰规律,结果表明大多数林窗是在50年以前形成的,这与罗浮栲种群数量场的震荡周期基本吻合。此外,罗浮栲群落在β值很小的时期,由于种群克服系统生态场“阻力”的潜能较小,需要采取人工保育措施,促进种群发展。
3) 森林生态系统是一个复杂的系统,一个群落只是森林生态系统中的一个“质点”,群落在受到生态系统对它的影响的同时,群落本身也对生态系统产生影响,本文基于这2种影响的综合效应展开研究,使研究结果反映了这2种影响的综合效应,符合客观实际。显然,本文的研究思路不同于早期群落生态场的研究观点,早期群落生态场研究主要着眼于群落对环境的影响,类似于对空间一个带电体,只注意它对周围的影响,这种思路用于城市或荒山、荒地中的群落生态场研究比较适合,如Penridge等(1988)对树冠光生态场的分布研究、蔺银鼎等(2006)对城市绿地空间结构对绿地生态场影响的研究等。
4) 本文主要是研究罗浮栲群落种群生态数量场,数量场不涉及场因子的矢量。如果要解释群落生态场中的物种发育、物种关系等具有矢量性质的问题,必须进行生态矢量场的研究,矢量生态场的研究更有利于深刻认识群落的发育、发展、演化等规律,为监测、调控和管理森林群落提供理论依据。
丛沛桐, 于景华, 赵则海, 等. 2001. 东灵山辽东栎林植物生态场研究. 植物研究, 21(1): 21-27. |
邓聚龙. 1985. 灰色系统. 北京: 国防工业出版社.
|
邓贤兰, 黄彩霞, 芦站根. 2005. 井冈山自然保护区栲树群落研究. 衡水学院学报, 7(1): 38-46. |
邓贤兰, 刘玉成, 吴杨. 2003. 井冈山自然保护区栲属群落优势种群的种间联结关系研究. 植物生态学报, 27(4): 531-536. DOI:10.3321/j.issn:1005-264X.2003.04.016 |
邓贤兰. 2003. 井冈山自然保护区栲属群落植物区系分析. 武汉植物学研究, 21(1): 61-65. DOI:10.3969/j.issn.2095-0837.2003.01.011 |
董振堂, 吕凤. 1989. 红松种群生态场特性函数初步分析. 植物研究, 9(4): 91-98. |
火树华, 康木生, 黄普华, 等. 1980. 树木学. 北京: 中国林业出版社.
|
蔺银鼎, 韩学孟, 武小刚, 等. 2006. 城市绿地空间结构对绿地生态场的影响. 生态学报, 26(10): 3340-3346. |
林英, 陆中光, 杨方西, 等. 1990. 井冈山自然保护区考察研究. 北京: 新华出版社.
|
刘金福, 洪伟, 李俊清, 等. 2003. 格氏栲林林窗自然干扰规律. 生态学报, 23(10): 192-197. |
刘仁林, 陈少凤. 2000. 井冈山大果马蹄荷林种群生态数量场的研究. 南昌大学学报, 24(2): 199-204. DOI:10.3969/j.issn.1006-0464.2000.02.021 |
谢树艺. 1985. 矢量分析与场论. 北京: 高等教育出版社.
|
王德利. 1994. 植物生态场导论. 长春: 吉林科学技术出版社.
|
王亚秋, 王德利. 2005. 改进的植物生态场模型与实例分析. 生态学报, 25(11): 2856-2861. |
邹春静, 韩士杰, 徐天铎. 1999. 生态场理论在沙地云杉种群更新中的应用. 生态学报, 19(1): 50-54. DOI:10.3321/j.issn:1000-0933.1999.01.007 |
Fabrig L. 1988. Nature of the ecological theories. Ecology Modeling, 43: 129-132. DOI:10.1016/0304-3800(88)90077-4 |
Gates. 1980. Biophysical Ecology. New York: Springer-Verlag.
|
Law R, Murrell D J, Dieckmann U. 2003. Population growth in space and time: Spatial logistic equations. Ecology, 84(1): 252-262. DOI:10.1890/0012-9658(2003)084[0252:PGISAT]2.0.CO;2 |
Maestre F T, Valladares F, Reynolds J. 2005. Is the change of plant-plant interactions with abiotic stress predictable? a meta-analysis of field results in arid environments. Ecology, 93: 748-757. DOI:10.1111/j.1365-2745.2005.01017.x |
Masaki H. 1999. A theoretical analysis and field evaluation of a light and temperature model of production by Ecklonia cava. Hydrobiologia, 398: 361-374. |
Penridge L K, Walker J. 1988. The crown-gap ratio (C) and crown cover: the field study. Ecology, 13: 109-120. |
Ronald E T. 1985. Distribution, abundance and reproductive success in the coral reef fish Acanthochromics polyacanthus. Ecology, 66(4): 1139-1150. DOI:10.2307/1939166 |
Sharpe P J H, Walker J, Penridge L K, et al. 1986. Spatial considerations in physiological models of tree growth. Tree Physiology, 2: 403-421. DOI:10.1093/treephys/2.1-2-3.403 |
Walker J, Crapper P E. 1989. Ecological field theory: the concept and field test. Vegetation, 83: 81-95. DOI:10.1007/BF00031682 |
Wildova′ R, Gough L, Herben T, et al. Architectural and growth traits differ in effects on performance of clonal plants: an analysis using a field-parameterized simulation model. Oikos, 116: 836-852. DOI:10.1111/oik.2007.116.issue-5 |