 2008, Vol. 44
2008, Vol. 44文章信息
- 张真, 李典谟.
- Zhang Zhen, Li Dianmo
- 马尾松毛虫暴发机制分析
- Approach to Outbreak Mechanism of Dendrolimus punctatus (Lepidoptera: Lasiocampidae)
- 林业科学, 2008, 44(1): 140-150.
- Scientia Silvae Sinicae, 2008, 44(1): 140-150.
-
文章历史
- 收稿日期:2006-09-21
-
作者相关文章



2. 中国科学院动物研究所 北京 100101
2. Institute of Zoology, Chinese Academy of Sciences Beijing 100101
从20世纪50年代开始我国就对马尾松毛虫(Dendrolimus punctatus)进行了大量研究,尤其是80年代以后,又进行了“六五”—“九五”攻关和自然科学重点基金等一系列研究计划,对马尾松毛虫的生物学特性、天敌、种群动态、发生规律和综合管理进行了大量研究(侯陶谦,1987;陈昌洁,1990),取得了很多成果。本文通过作者的研究,结合以往的研究成果,探讨马尾松毛虫的发生机制,以利于更加深入地认识马尾松毛虫的发生规律,更加有效地对马尾松毛虫进行持续管理。
害虫的暴发从生态学角度看是特定种群的数量在相对短的一段时间内急剧增加的现象。Berryman(1987)将害虫暴发的原因总结为如下的假说:1)物理环境条件的急剧变化,属于这类原因的包括太阳黑子理论(Elton,1924)、气候释放论(Greenbank,1956)、环境理论(Andrewartha et al., 1984);2)种群个体内的遗传(Chitty,1971)和生理(Wellington,1960; Christian et al., 1971)变化;3)植物与植食昆虫之间或捕食者和猎物之间关系变化(Lotka,1925;Nicholson et al., 1935);4)环境胁迫引起寄主植物质和量的变化(White,1978;Mattson et al., 1975);5)容易引起暴发的特定生活史对策(Southwood et al., 1976;Rhoades,1985;Nylin,2001);6)害虫种群摆脱了天敌的调节作用(Holling,1965;Morris,1963;Takahasi,1964);7)种群间的互惠作用超过了寄主的防卫作用(Berryman,1982a;1982b)。
以上假设的提出都是根据各自研究的对象和方法得出的结论,由于生命系统本身的多样性和环境变化的多样性,以上假设分别适应于一些特定的对象,对于有些对象的暴发原因可能不能用一种假设来解释,各假设之间还有一定的联系和相互重叠。Berryman(1987)通过对理论模型的分析进一步将其归结为4个方面的变化:1)有利的环境;2)更多的高产卵量基因型;3)更多抗天敌基因型;4)适应型和协同型的增加。另外,种群的迁入和短期的环境和遗传波动,也会引起害虫的暴发,这种波动只需要将种群提高到暴发域值,超过这个域值种群的暴发就会自动地进行,即使环境和遗传条件返回到正常水平也不能阻止暴发过程。Berryman(1987)还认为有3个重要的特征决定了种群暴发的时空动态:1)有的暴发是自我维持型(self-perpetuating),暴发一旦触发,就会在时空上不断发展,这种类型受较高密度下的正反馈控制,被称作“突发性(eruptive)暴发”;2)暴发完全依赖于外部环境和内部遗传条件,其起伏波动随驱动因子而变化,被称作“梯度(gradient)暴发”;3)种群时空动态受具时滞的负反馈所决定,时滞越长围绕平衡点的波动越剧烈,时滞通常由天敌和寄主的时间依赖性反应而产生,物理环境和基因频率的变化使种群交替出现维持、增长和下降。即周期型暴发。他还根据这3个特征组合成8种暴发类型。
近年的研究较多地从分子生态学(Hanski et al., 2006;Watt et al., 2003;Wheat et al., 2006)、寄主-害虫-天敌的3重营养关系(Denno,2002)、景观生态学(Cappuccino et al., 1998;Radeloff et al., 2000;Johnson et al., 2004)和气候变化(Zhang et al., 1999;Allan et al., 2002;Battisti et al., 2006)的角度研究害虫的暴发机制,进一步补充和丰富了以上假说,但还没有跳出这些假说的内容。
总之,害虫的暴发受害虫本身的遗传特征、寄主、天敌和物理环境因子及其相互关系所确定,不同的种类,处于不同的环境条件,会具有不同的暴发特点和暴发机制,马尾松毛虫暴发的机制是什么呢?以下将在分析马尾松毛虫的特性及其与各种环境因子的相互关系的基础上,讨论其暴发机制。
1 马尾松毛虫暴发的原因自然界有很多昆虫,真正能够暴发成灾的是很少的种类。一种昆虫能暴发成灾,与它本生的特性和所处的环境有关,以下就从这2个方面分析其暴发的原因。
1.1 马尾松毛虫的暴发特性我国共有松毛虫27种,但在这27种松毛虫中只有6种经常成灾,而马尾松毛虫危害成灾的程度列为第1,其他种类的顺序为落叶松毛虫(D. superans)、油松毛虫、赤松毛虫、云南松毛虫、思茅松毛虫。
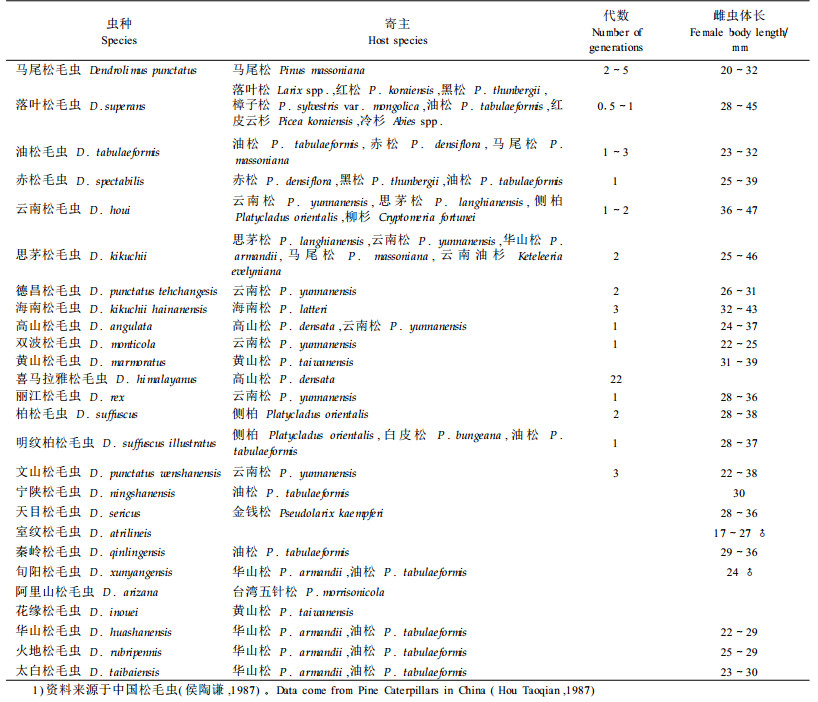
已有很多专家研究了暴发种的特性,尤其是同一类昆虫,为什么有的种经常暴发成灾,而有的种数量一直保持较低密度,从不引起明显的危害?虽然寄主、天敌、气候等因子都与害虫的暴发有关,但在相似的生境条件下有的种能暴发,有的种却不能暴发,这些暴发种必然具有与非暴发种不同的生活史特点或对气候、寄主、天敌变化具有不同的反应(Hunter,1991)。Watt(1964)总结加拿大大型鳞翅目昆虫后认为波动较大的种类具有较宽的寄主范围。然而,Redfearn等(1987)却有与之相反的发现,他们的分析结果表明种群变动幅度与寄主范围成反比或无关。Spitzer(1984)发现夜蛾科昆虫的种群变动大的种类具有较大的产卵量。Hunter(1991)对北方阔叶树的暴发种和非暴发种的生活史特性进行比较研究认为多数的暴发种为春季危害的种,或具有聚集特性的夏季危害种,暴发种的食性范围较宽,产卵量较高。这是因为春季危害的种易受春天多变气候的影响而波动,聚集性种类的不同聚块大小直接影响到对捕食性天敌抵抗能力。高的产卵量和较宽的食性范围有利于在条件适合的情况下迅速增长。
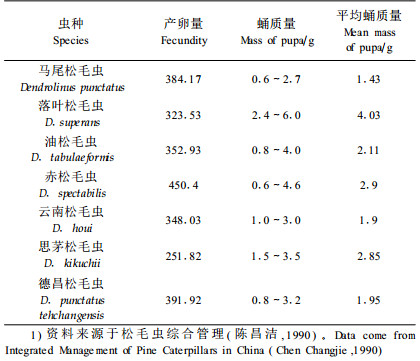
由于缺乏松毛虫非暴发种的资料,很难比较暴发种和非暴发种的特性,表 1、2列出了已有松毛虫形态和生物学特性的数据。从松毛虫的生物学特性分析,多数暴发种的寄主范围较广,产卵量较高,是暴发成灾的一个重要因素,但马尾松毛虫的寄主范围不广,产卵量也不是最高的,但成灾的程度却第一,说明还有其他因素导致马尾松毛虫的成灾。
|
|
|
|
寄主的生理、生化和物理特性特点和动态变化不但会直接影响害虫的发生和动态,还会间接地通过天敌的寄主和直接地影响天敌,这种“自下而上的因子”(bottom up factors)与天敌的“自上而下的因子(top down factors)”具有复杂的相互关系,这种关系作用的结果决定了害虫种群的状态(Denno,2002)。寄主的种类和状态直接影响马尾松毛虫的发生和动态,通常生长健壮的松树抗性较强。广西玉林地区林业科学研究院的研究表明,感虫的马尾松的游离氨基酸总量比抗虫植株增加32%~58%。游离氨基酸总量的增加,是衰老后的特征性变化,说明蛋白质合成减少而降解增强。
马尾松毛虫在一般情况下喜吃老针叶而不喜吃幼嫩针叶,由于老针叶水分少,酸的含量亦较低,同时纤维素、可溶性碳水化合物及蛋白质含量较高,营养物质较丰富,有助于松毛虫的生长发育(侯陶谦,1987)。
马尾松受害程度越高,马尾松毛虫的产卵量、蛹质量、雌性比、繁殖量越低,死亡率越高,原因主要是针叶中所含化学营养物质不同。松树受松毛虫中等程度危害后,棕榈酸、生物碱、单宁等不利于松毛虫生长的物质增加,而蛋白质、β蒎烯水溶性总糖、C/N等营养物和吸引松毛虫的物质减少(侯陶谦,1987;戈峰等,2002)。但李海林等(2005)的研究表明幼虫的取食量、排粪量、蛹质量、对食物的消耗和利用指数、化蛹率和生殖力以取食未受害松树(针叶损失0)的最高,重度受害(针叶损失75%)次之,轻度受害(针叶损失25%)再次,中度危害(针叶损失50%)最低,而发育历期和死亡率则相反。这可能由于当针叶损失在25%时,针叶内产生的化学物质变化,植株处于警戒状态;当针叶损失50%时,针叶内的次生代谢物质的含量增加到最高点,引发松树产生诱导化学防御的能力也最强;在针叶损失75%的马尾松针叶内,次生代谢物质和营养含量与针叶损失25%时差异不大(王燕,2001;章康华2002)。
不同种的寄主对马尾松毛虫也有不同的影响。用不同树种饲养马尾松毛虫,其喜好程度为马尾松(Pinus massoniana)最大、湿地松(P. elliottii)次之,火炬松(P. taeda)最小,可见马尾松是马尾松毛虫最适宜的寄主(安徽林科所,1976;苏星等,1983;李文杰等,1983;刘兴平等,2003)。国外松针叶数比马尾松多11.5%~47.9%,平均每根针叶长度比马尾松长23.7%~37.3%。同样受到马尾松毛虫危害后,3种国外松的生长量较马尾松快得多,表明其耐害性比马尾松强。国外松抗松毛虫通过机械屏障和化学屏障2个方面。马尾松针叶只有表皮细胞是厚壁细胞,皮下细胞均为薄壁细胞;而3种国外松除表皮外一部分皮下细胞也为厚壁细胞。国外松的维生素B6含量、挥发物含量比马尾松低,而这2个因子是影响马尾松毛虫化蛹、羽化和周限增长率第1和第2位的正相关重要因子(邹运鼎等,1990)。马尾松毛虫幼虫在第1代发生期取食马尾松老叶、新叶和湿地松老叶的幼虫成活率分别为44%、32.7%、4%,而取食湿地松新叶的幼虫全部死亡。用增加了外源β-蒎烯的松针饲养的马尾松毛虫幼虫与未增加外源β-蒎烯饲养的幼虫相比,死亡率明显提高;松树中β-蒎烯含量的增加与幼虫成活率呈显著负相关(何中等,1997)。
寄主不但影响松毛虫,也间接地影响天敌,在对松毛虫赤眼蜂(Trichogramma dendrolimi)雄蜂寄主栖息地定向行为的研究中发现,来自马尾松松针的气味能刺激松毛虫赤眼蜂雄蜂加强在气味区的搜索活动,但仅有松针气味尚不能明显地将雄蜂滞留在气味区内;同种雌蜂的气味对雄蜂虽然有明显的引诱作用,但对雄蜂的滞留也无明显的影响;当松针加上雌蜂时,气味区的雄蜂头数明显地增加(邱中良等,1995)。把马尾松松针和玉米叶同时提供给松毛虫赤眼蜂雌蜂时,雌蜂明显选择马尾松针叶(沈伯均等,1991)。
马尾松毛虫在混交林中的发生轻于纯林,复层林轻于单层林,8~20年生的松树最易受害,而天敌的种群数量和种类与之相反,混交林中的多于纯林,复层林多于单层林。马尾松毛虫在马尾松纯林中发生量多,为害最严重,暴发周期最短;在与杉木等混交的马尾松林中发生次之;而在与阔叶树混交的马尾松林中平均每株虫口数少,发生与危害率及有虫株率均较其他林型小,不易引起松毛虫暴发为害(侯陶谦,1987;张执中等,1990)。
马尾松林景观结构对马尾松毛虫的发生有明显影响,从斑块水平分析,马尾松林斑块面积和形状指数与马尾松毛虫的发生率和发生面积成明显的正相关。从景观水平分析,平均斑块指数和斑块丰富度指数与马尾松毛虫的发生率成明显的正相关,该结果说明大面积马尾松林的存在有利于马尾松毛虫种群的繁衍,边缘效益对马尾松毛虫的作用明显,林缘比例越大,马尾松毛虫的发生越严重,林缘的小气候条件对松毛虫的发生十分有利(张真,2003)。
从以上分析可以看出,马尾松是马尾松毛虫的适宜寄主,所以大面积马尾松的存在,有利于马尾松毛虫的发生,是导致马尾致松毛虫暴发的重要因素。
2 天敌对马尾松毛虫动态的影响由于马尾松毛虫广泛分布于我国南方的15个省区,所以天敌种类也很丰富,共有天敌399种(陈昌洁,1990)。
卵期寄生性天敌中,重要有松毛虫赤眼蜂、松毛虫黑卵蜂(Telenomus dendrolimus)和白跗平腹小蜂(Pseudanastatus albitarsis),幼虫期的松毛虫内茧蜂(Rogas dendrolimi)、蛹期的大腿小蜂(Brachymeria donganensis)和姬蜂(Casinaria nigripes)等,寄生蝇主要有蚕饰腹寄蝇(Blepharipa zebina)、伞裙追寄蝇(Exorista civilis)、红尾追寄蝇(E. xanthaspis),其中蚕饰腹寄蝇多为优势种,寄生率的情况基本与卵期天敌相似,以低丘陵的纯林松毛虫猖獗后期、混交林和封山区寄生率较高(陈昌洁,1990)。寄生性天敌的寄生率的高低与林型和发生阶段有关,一般高寄生率出现在混交林、灌木茂密、植被丰富的林地和纯林猖獗后期,寄生性天敌虽然在一定条件下能达到很高的寄生率,但在松毛虫平均总死亡率中的作用不大,如据浙江1979—1981和1984—1985年马尾松毛虫生命表统计,寄生天敌所致死亡率仅占总死亡率的0.77%(0.02~2.89)(陈昌洁,1990;柴希明,1995)。
捕食性天敌的种类很多,作用较大的有蚂蚁、蜘蛛、螳螂类、猎蝽类、鸟类(陈昌洁,1990),是幼虫期的重要天敌,捕食性天敌除直接捕食的作用以外,更重要的是引起松毛虫小幼虫的分散、扩散、吐丝下垂,由此造成大量死亡。柴希明于1979—1980年对越冬代松毛虫1龄幼虫的直接捕食只占该期总死亡率的1%。捕食作用引起的死亡率往往占总死亡率的49.2%~86.63%。尤其在松毛虫虫口密度较低的潜伏期时,捕食天敌为主要的死亡因子。捕食性天敌多为多食性,由于食量有限,虫口密度低时,捕食率较高,虫口密度高时,总捕食率反而较低,所以一般情况下捕食性天敌对松毛虫的作用为负密度相关(柴希民,1995;石根生等,1998)
感染马尾松毛虫的微生物主要有白僵菌(Beauveria spp.)、苏云金杆菌(Bacillus thuringiensis)、病毒(松毛虫CPV),在自然界有时发生自然流行(侯陶谦,1987),昆虫致病菌的流行与环境、松毛虫、松毛虫的寄主和昆虫群落结构都有关系,松毛虫密度和林间白僵菌的量是马尾松毛虫白僵病林间流行的机制中最重要的决定性因素,郁闭度是森林小气候的一个重要方面,三者结合才能使白僵菌的流行成为可能(高丙利等,1994)。白僵菌的流行多发生在越冬代,尤其是在连续几天阴雨的条件下,白僵菌感染率最高,可达97%~100%(柴希民,1995;梁修山等,1999)。
大量的研究表明,植被丰富,中间寄主多的林地,天敌的种类和数量也多,封山育林区天敌的种类和数量都大于非封山区。而天敌的种群数量和种类混交林中的多于纯林,复层林多于单层林(侯陶谦,1987;张执中等,1990;任立宗等,1988;任立宗,1990)。
马尾松毛虫的发生可分为潜伏期、上升期、猖獗期和衰退期。马尾松毛虫潜伏期捕食是主要的死亡因子,卵期鸟类的捕食达77.1%,第1、2、3代幼虫期蜘蛛、蚂蚁、捕食蝽等的捕食率分别达到84.3%、79.15%和99.5%,茧期鸟类捕食率达95.9%,成虫期鸟类的捕食率最高100%,最低24%(柴希明等,1992)。在马尾松毛虫上升期,天敌的种类和数量及整个群落的多样性随松毛虫的增加而增加,如果环境条件较好,能在一定程度上抑制马尾松毛虫的发生(如在偶灾区),在常灾区天敌数量的上升跟不上松毛虫的上升,松毛虫将大暴发(张真等,1998)。在松毛虫的猖獗期和下降期,寄生性天敌和松毛虫病菌的寄生率增加,同时由于食物短缺,产卵量和雌性比减少,使松毛虫的数量下降到低水平。
从以上分析中可以认为天敌,尤其是捕食性天敌在维持马尾松毛虫种群于低水平起到重要作用,但如果群落的整体抑制作用不够强的情况下,在马尾松毛虫迅速上升期不能起到有效的抑制作用,导致松毛虫的暴发,在暴发的后期寄生性天敌和病菌在松毛虫种群的下降期起重要作用。
3 气候因子和立地条件对马尾松毛虫动态的影响气候因子对害虫的影响是多方面的,不但会直接影响害虫的生长发育和存活,还会影响寄主生理生化状态、天敌及群落中其他昆虫,从而间接地影响害虫的种群动态。
马尾松毛虫发育需要一定的积温,在马尾松毛虫分布区,从北到南随年平均温度升高,马尾松毛虫1年内完成的代数增加,如河南和安徽北部以2代为主,湖北、湖南、安徽南部等地1年发生2~3代,广东、广西、福建等地1年发生3~4代,海南岛每年可以发生5代(侯陶谦,1987)。
各地不同年份之间的世代分化比有时变化很大,在马尾松毛虫世代分化以前温度越高,发育越快,在滞育的临界光周期出现之前发育的幼龄幼虫越多,滞育幼虫就越少,分化比就越大。温变系数也会影响马尾松毛虫的发育,在平均温度相差不大的情况下,温度变动大,幼虫发育慢,温度变动小,幼虫发育快(张宏,1993)。马尾松毛虫幼虫死亡率在长光周处理内低温组为25.55%,常温组为55.13%,高温组为58.23%;在短光周处理内低温组为32.67%,常温组为64.67%,高温组为94.32%,说明高温是影响马尾松毛虫低龄幼虫死亡的主要原因(何忠等,2006)。
马尾松毛虫在长日照条件下,生长快,发育良好,短日照下发生滞育,光周期是产生世代分化的关键因子,恒温和变温条件下,临界光周期分别为13 h 33 min和13 h 46 min(李兆麟等,1995)。光周期的感受主要是幼龄幼虫期,但马尾松毛虫的整个幼虫期都处在短光照周期条件下,才能维持幼虫的滞育,滞育强度较弱,解除滞育的条件也不严格,属于浅滞育类型。但在第2代幼虫孵出后20 d内如遇连续高温环境,即使在短光周条件下,幼虫也不会滞育(何忠等,2006)。所以,温度、虫口密度、被害率等因子对马尾松毛虫的滞育和世代分化也有一定的影响(李兆麟等,1993;1994;1995;何庆全等,1995;王常禄等,1995)。
松毛虫的卵、幼虫和蛹如长期处于相对湿度小于75%的条件下,不能保持正常的生长发育,而雨量多少直接影响温湿度的高低,马尾松毛虫发生地区,4—5月间阴雨连绵,有利于白僵菌等微生物的繁殖,增加松毛虫的死亡。在2~3代发生区5月和8月的降水总量大,容易引起马尾松毛虫的大发生,这是因为这时正是卵孵化和1~3龄幼虫期,此时气温比较稳定,在适温范围内,降水量增多,湿度必然增大,有利于卵的孵化和幼龄幼虫的存活。但短时间暴雨能将幼虫从树上冲刷下来,而长时间连绵雨会促进幼虫的结茧、化蛹和成虫羽化(侯陶谦,1987)。李振华(1997)研究发现全年雨日及11月雨日越少,当年马尾松毛虫发生量越大,次年越冬代恢复取食危害面积及程度也越大。干旱只与下一年的第1代马尾松毛虫的发生有显著正相关。说明干旱对松毛虫时间动态的影响具有延迟作用,干旱不仅影响当代松毛虫的动态,更重要的是通过影响寄主,从而影响到以后世代的松毛虫动态(张真等,2002)。
风向和风力主要影响松毛虫的扩散方向和传播面积,幼虫和成虫常借风力扩散,形成定向迁移,但强台风和暴雨配合,常造成松毛虫大量死亡。
马尾松毛虫一般阳坡高于阴坡,西坡高于东坡,特别是虫源地和初发期,特征期更明显。而且阳坡世代分化也明显高于阴坡。坡度的大小可以使坡向的影响加强或减弱。坡度大的山地,山峦起伏,小生境交错重叠,不容易发生松毛虫虫灾,即使发生也易被天敌和微生物所控制,一般林缘比林内严重(彭建文,1990)。本文在松林景观结构对马尾松毛虫发生影响的分析中也进一步说明松林斑块形状指数越大,即林缘比例越大,松毛虫发生越严重。这说明阳光充足高温有利于马尾松毛虫的发生。
海拔对马尾松毛虫的影响非常明显,一般情况下,500~700 m以上的山区为无灾区,400~500 m的半山区为偶灾区,低海拔的丘陵地区为常灾区(侯陶谦,1987;张执中等,1990)。主要由于高海拔地区低温多湿、日照短,不适于马尾松毛虫的生长发育。
4 马尾松毛虫种群时空动态及发生规律 4.1 空间分布及动态频次检验法对马尾松毛虫越冬幼虫空间分布型的研究表明,越冬幼虫呈负二项分布,为聚集空间格局(王淑芬等,1983)。分布型指数法研究结果显示,其卵块呈随机分布,小幼虫、大幼虫及茧呈聚集分布。聚集程度为小幼虫最高,茧最低,幼虫及茧的的基本成分是疏散的个体群(李天生等,1981)。地统计学方法研究不同松林马尾松毛虫蛹及其寄生天敌的空间格局结果表明:在混交林中,松毛虫蛹为聚集分布,空间结构模型为球型,天敌的分布为随机分布或均匀分布,空间结构模型为直线;而在纯林中低密度时,松毛虫蛹的空间结构为随机分布或均匀分布,空间结构模型为直线,高密度时为聚集分布,空间结构模型为指数,但不管是低密度还是高密度,天敌均为随机分布或均匀分布,空间结构模型都为直线(石根生等,1997)。柴希明(1995)研究了40块标准地1972—1974年间马尾松毛虫重危害松林的空间动态,他将松毛虫的危害过程描述为“蚕食—镶嵌—连片”。
4.2 不同发生阶段的动态特点马尾松毛虫的发生有一定的周期性,一般可分为潜伏期、上升期、猖獗期和衰退期。马尾松毛虫不同发生阶段的动态特点是不同的,在密度很低的潜伏期,松毛虫的产卵量、雌性比维持一定的水平,而死亡率很高,死亡的主要原因是捕食性天敌;而在松毛虫的上升期,松毛虫的产卵量和雌性比成较大幅度增加,虫口密度迅速猛增,种群的聚集程度增加;到猖獗期,松毛虫开始扩散,天敌寄生率开始上升,由于食物短缺,产卵量、性比开始下降;到衰退期,松毛虫的羽化率、雌性比降到很低的水平,而寄生率上升到高水平,松毛虫密度迅速下降到非常低的水平。
4.3 暴发类型和周期性大量发生的历史记录表明,马尾松毛虫的暴发具有一定的周期性,一般每隔3~5年大暴发1次,根据成灾的程度和频次可将其划分为常灾区、偶灾区和无灾区(侯陶谦,1987)。柴希明(1995)将马尾松毛虫的暴发类型分为4种类型:1)渐进发生型 松毛虫在马尾松林中在低水平上波动,大发生前松毛虫数量逐渐上升,经过4~6个世代导致大发生2)暴发发生型 在大发生前松毛虫有一定密度,数量在低密度范围内波动,当环境条件适宜时,松毛虫数量急增,暴发成灾,一般为虫源地;3)扩散发生型 原来松毛虫虫口密度极少,由暴发发生型林地松毛虫飞来产卵,使松毛虫数量突然增加;4)低密度稳定发生型 松毛虫密度低,数量变动幅度小,松林被害轻微。松毛虫常灾区多为暴发发生型,偶灾区一般为其他3种类型。而暴发发生型是松毛虫的典型发生类型。
5 松毛虫发生与发展机制分析 5.1 引起松毛虫快速暴发的机制大青山实验林场马尾松毛虫一个暴发周期的虫口密度变化和分布如图 1。

|
图 1 马尾松毛虫1个暴发周期虫口密度的分布 Figure 1 Density distribution of MPC during an outbreak period |
图 1说明常灾区松毛虫不仅虫口密度变动大,而且变动过程为从低密度迅速上升到高密度,中间没有过渡,而下降迅速,所以虫口密度分布成双峰型,且低密度频率高,高密度频率低。而偶灾区和无灾区变动幅度小,频率分布较为均匀。马尾松毛虫在常灾区为什么能快速暴发,而在潜伏期虫口密度却很低,这种短期内大幅度波动的机制是什么?本文及前人的研究认为:由于潜伏期捕食性天敌的作用很强,使马尾松毛虫处于低密度,如果此时处于暴发后期,松树还处于恢复期,无论如何松毛虫都不会上升,种群维持在低水平,这种状态持续一段时间之后(一般1~2年),松树的状态逐渐得到恢复,这时松毛虫已经具备了暴发的物质基础,一旦出现有利的环境因子,松毛虫种群密度将会有一定程度的上升,如果此时系统的多样性程度不高,不能有效地抑制松毛虫的上升,松毛虫就会摆脱天敌的控制而迅速上升,暴发成灾。Berryman(1987)指出迅速上升阶段必然存在正密度相关(positive density dependant)的作用,即此阶段种群的个体之间存在“互相协助(cooperate with one another)”的关系而不是互相竞争,随密度增加生殖率和存活率增加,马尾松毛虫是否在上升期存在正密度相关呢?由于马尾松毛虫在常灾区低密度时,空间结构为随机分布或均匀分布,高密度时为聚集分布,但不管是低密度还是高密度,天敌均为随机分布或均匀分布(石根生等,1997),所以在松毛虫种群的上升期,随密度增加,聚集度增加,而天敌的聚集度不能增加,聚集度增加的结果,对天敌的抵抗力增加,存活率增加,所以天敌的作用随密度增加而降低,从而导致正密度效益。石根生等(1998)研究捕食性天敌10种对马尾松毛虫低龄幼虫的捕食作用的结果表明斜纹猫蛛(Oxyopes sertatus)、细纹猫蛛(O. macilentus)、交迭圆蛛(Araneus alternidens)和大刀螳螂(Tenodera aridifolia)4种为Holling Ⅲ型功能反应,而其他几种与密度关系不显著。说明天敌的捕食率随松毛虫密度增加到一定程度后就随密度增加而迅速下降,或者任何密度下都只捕食一定数量。对黄荫规(1994)研究中的数据分析,发现了马尾松毛虫天敌数量与松毛虫数量成负相关。柴希民(1995)在鸟类对松毛虫捕食率的研究中也证实了正密度制约的作用。
在偶灾区和无灾区由于系统的多样性较高,抑制作用较强,使马尾松毛虫不易摆脱天敌的控制而进入正密度制约作用产生的快速增长,而只是有一定幅度的增长。
5.2 松毛虫快速下降的机制对马尾松毛虫时间序列的自相关分析表明,其动态为稳定平衡类型,暴发后很快回到低密度的水平,具有很强恢复力(张真等,2002)。为什么松毛虫暴发以后又会很快下降呢?一方面由于松毛虫对松树的诱导抗性和严重失叶,在受害后松树的抗虫成分增加,使松毛虫的雌性比降低,产卵率降低(戈峰等,2002);另一方面寄生性天敌和病菌的感染率增加。下降到一定密度以下后,由于天敌的作用,维持在很低的密度。
5.3 松毛虫从低密度到高密度变化的启动因子马尾松毛虫的暴发虽然有一定的周期性,但并不成严格的周期性,说明其何时暴发也存在一定的不确定性,需要有一定的启动因子。那么松毛虫怎么从低密度下的负密度制约转化为较高密度下的正密度制约呢?一是异常的气候条件,如高温(引起马尾松毛虫发育进度和生活史特性的变化)和干旱(寄主植物质和量的变化)(张真等,2002);另一种可能是环境噪音(noise)引起种群的混沌动态(chaotic dynamics)(张真等,2002;2003)。即Berryman(1987)所列的假说1、4和5。
5.4 生态系统对松毛虫的抵抗机制生态系统的多样性,可以降低松毛虫的发生程度和发生率。具体来说,多样性包括几个方面的内容:1)景观结构的多样性,单一成片的马尾松林中马尾松毛虫最易暴发成灾;2)植被的多样性,植被多样性高,导致昆虫群落及其天敌的多样性,从而抑制松毛虫的发生;3)食叶类群昆虫的多样性,食叶类群的多样性导致同一营养层的竞争加剧,从而抑制了松毛虫的发生。另一方面上升期在多样性较高的情况下,虫口密度上升幅度较小,松树失叶不超过50%,由于松毛虫对松树的诱导抗性的作用,使虫口密度下降。另外对马尾松毛虫不利的气候因子导致松毛虫不能成灾,如高海拔地区。
5.5 松毛虫的周期性暴发的机制一般认为马尾松毛虫暴发具有周期性,但这种周期性不是数学意义的严格的周期性,对马尾松毛虫时间序列的自相关函数分析表明其周期性不显著,而且空间尺度越大,周期性越不显著,所以只能说它的暴发成大致的周期性(张真等,2002)。造成周期性的原因是具时滞的密度制约,如松树抗性反应的时滞(戈峰等,2002)和一些寄生性天敌和致病菌作用的时滞。
总之,马尾松毛虫的暴发过程具有周期性和突发性相结合的特点,不同环境条件下,即不同的发生区,发生类型不一样,常灾区是典型的暴发发生类型,偶灾区和无灾区一般为渐进发生型、扩散发生型和低密度稳定发生型。暴发的原因一方面因为马尾松毛虫具备了成为暴发种的遗传特性,另一方面是由于大面积马尾松纯林的存在。暴发的启动因子为高温干旱或由环境噪音引起的混沌动态,暴发的上升期为正密度制约作用驱使,下降期和潜伏期主要是负密度制约的作用。图 2表示了松毛虫暴发的机制。

|
图 2 马尾松毛虫暴发过程及其机制 Figure 2 Outbreak process and mechanism of MPC |
综上所述,马尾松毛虫的暴发过程具有周期性和突发性相结合的特点,不同环境条件下,即不同的发生区,发生类型不一样,常灾区是典型的暴发发生类型,偶灾区和无灾区一般为渐进发生型、扩散发生型和低密度稳定发生型。暴发的原因一方面是马尾松毛虫具备了成为暴发种的遗传特性,另一方面是大面积马尾松纯林的存在。暴发的启动因子为高温干旱或由环境噪音引起的混沌动态,暴发的上升期为正密度相关作用驱使,下降期和潜伏其主要是由松树诱导抗性、严重失叶和天敌所引起的负密度相关的作用。松林和松林节肢动物群落结构的多样性是抑制马尾松毛虫暴发的主要因素。
对像马尾松毛虫这样的历史性大害虫,已积累的基本素材较多,对它的基本生活史、生物学、生态学、天敌和发生规律有较多的了解,在实践中也积累了很多有效的综合管理技术,但对其发生机制和深层次的规律仍不十分清楚,是进一步进行深入研究害虫暴发机制和生态管理的合适对象,本文的研究只是这方面的一点探索,很多地方证据还不够充分,今后还需在基因、种群、群落和景观不同层次上在如下几个方面进行深入研究:
1) 目前对马尾松毛虫的遗传结构和分子生态学了解还非常少,研究松毛虫暴发过程中基因的调节机制极其作用,从这方面研究松毛虫的暴发机制能使我们了解其暴发的本质。2)从种群动态的角度应深入研究种群暴发过程中从低密度到高密度的突变规律及这种突变的产生和控制机制。3)研究松毛虫动态变化过程中性信息素活性和含量变化及松毛虫对性信息素的敏感性,探明这些变化在松毛虫暴发过程中的作用。4)从生理生化和化学生态学的角度研究松林生态系统的化学信息网极其松林群落组成结构与节肢动物群落组成结构之间的关系和动态调节过程及机理,揭示多样性与稳定性关系的本质。5)进一步研究在松毛虫不同密度下及暴发过程中,天敌种类和作用的差异,深入了解天敌对松毛虫种群的调节机制。6)以往对马尾松毛虫的研究较少从时空动态和大空间尺度上进行,本研究只是从景观生态学的角度进行了初步的研究,今后应从这个方向进行深入的研究,包括不同时空尺度上景观结构、多样性及其动态对昆虫群落动态和害虫、天敌之间关系的影响,如何通过景观结构调整,增加系统的多样性和稳定性。7)在深入研究马尾松毛虫的动态发生规律和暴发机制的基础上,应加强对马尾松毛虫持续控制技术的研究,这种技术除林分结构改造外,应着重通过监测手段,准确掌握松毛虫的上升期,确定生态管理指标,在上升期采取有效的生物防治手段进行防治,避免马尾松毛虫灾害的发生。
柴希明, 何志华, 吴正东. 1992. 潜伏期马尾松毛虫死亡因子分析. 林业科学, 28(2): 184-187. |
柴希明. 1995. 马尾松毛虫种群动态研究. 浙江林业科技: 1-81. |
陈昌洁. 1990. 中国松毛虫综合管理. 北京: 中国林业出版社.
|
高丙利, 吴东亮, 王淑芬, 等. 1994. 马尾松毛虫白僵菌流行在空间轴上的动态变化. 中南林学院学报, 14(2): 102-108. |
戈峰, 李镇宇, 谢映平, 等. 2002. 我国主要松树诱导抗虫性的一些规律比较. 北京林业大学学报, 24(3): 61-65. DOI:10.3321/j.issn:1000-1522.2002.03.013 |
何庆全. 1995. 马尾松毛虫2、3代分化规律探讨. 浙江林业科技, 15(2): 38-41. |
何忠, 韩瑞东, 刘向辉, 等. 2006. 环境温度对马尾松毛虫发育与存活的影响. 应用生态学报, 17(3): 483-488. DOI:10.3321/j.issn:1001-9332.2006.03.025 |
何忠, 曹红珍, 曾菊平, 等. 2007. 马尾松毛虫对马尾松和湿地松的选择与适应研究. 昆虫学报, 50(2): 125-135. DOI:10.3321/j.issn:0454-6296.2007.02.006 |
候陶谦. 1987. 中国松毛虫. 北京: 科学出版社.
|
黄荫规. 1994. 马尾松毛虫食物链及其应用研究. 广西林业科学, 2: 55-56. |
李天生, 柴希民, 吴征东. 1981. 马尾松毛虫(Dendrolimus punctatus)的空间分布型及其在实践上的应用. 林业科学, 17(4): 343-350. |
李文杰. 1983. 五种松树对马尾松毛虫抗性的初步研究. 安徽林业科技, (2): 15-19. |
李兆麟, 何忠, 贾凤友. 1993. 马尾松毛虫幼虫发育的光周效应. 林业科学研究, 6(3): 276-281. DOI:10.3321/j.issn:1001-1498.1993.03.008 |
李兆麟, 何忠, 贾凤友. 1995. 温周对马尾松毛虫的光周期反应的影响. 林业科学研究, 8(8): 413-416. |
李兆麟, 贾凤友, 侯无危, 等. 1994. 马尾松毛虫的光周期反应. 昆虫学报, 37(1): 31-37. |
李振华. 1997. 雨日对马尾松毛虫诱发作用的初探. 江西林业科技, (5): 32-33. |
李海林, 李镇宇, 张丽丽. 2005. 取食不同受害程度的马尾松对马尾松毛虫种群数量的影响. 中国森林昆虫, 24(6): 1-5. |
刘兴平, 王国红, 刘向辉, 等. 2003. 不同林相的松针对马尾松毛虫生长发育的影响. 昆虫学报, 46(6): 720-726. DOI:10.3321/j.issn:0454-6296.2003.06.009 |
梁修山, 章立新, 徐光宇. 1999. 林分因子和气候对松毛虫白僵病流行的影响. 森林病虫通讯, (4): 4-7. |
邱中良, 周建中, 黄娟, 等. 1995. 松毛虫赤眼蜂雄峰寄主栖息地定向行为的研究. 昆虫天敌, 17(1): 1-6. |
任立宗, 王淑芬. 1988. 马尾松昆虫群落及时空结构的研究. 林业科学研究, 1(4): 397-403. |
任立宗. 1990. 不同类型区天敌资源及其差异//陈昌洁. 松毛虫综合管理. 北京: 中国林业出版社, 119-124.
|
沈伯均, 邱鸿贵, 邱中良, 等. 1991. 松毛虫赤眼蜂对不同植物的行为反应. 昆虫知识, 28(6): 359-361. |
石根生, 李典谟. 1997. 不同松林马尾松毛虫蛹及其寄生天敌群子的空间格局分析. 生态学报, 17(4): 386-392. |
石根生, 李典谟. 1998. 马尾松毛虫幼虫的捕食天敌及其捕食作用的研究. 昆虫知识, 35(6): 336-340. |
苏星, 仪向东, 邓常发. 1983. 几种松树对马尾松毛虫抗性的初步研究. 林业科学, 19(增): 49-57. |
王常禄, 汪国华, 何庆全, 等. 1995. 马尾松毛虫的二、三代分化规律的研究. 林业科学, 31(6): 559-564. |
王国红, 刘兴平, 戈峰, 等. 2005. 不同松林内马尾松毛虫种群动态的特征. 生态学杂志, 24(4): 355-359. DOI:10.3321/j.issn:1000-4890.2005.04.001 |
王淑芬, 李去惑, 陈纪文, 等. 1983. 马尾松毛虫空间分布型及其抽样技术研究. 中南林学院学报, (3): 28-42. |
王燕, 戈峰, 李镇宇. 2001. 马尾松诱导化学物质变化的时空动态. 生态学报, 21(8): 1256-1261. DOI:10.3321/j.issn:1000-0933.2001.08.007 |
章康华, 陈春平, 刘兴平, 等. 2002. 马尾松诱导抗性对马尾松毛虫幼虫生长发育的影响. 昆虫知识, 39(4): 292-296. DOI:10.3969/j.issn.0452-8255.2002.04.013 |
张宏. 1993. 温变系数对马尾松毛虫发育关系的研究. 林业科学, 29(5): 468-471. |
张真, 李典谟, 张培仪, 等. 2003. 混沌的检测及其对种群生态学的意义. 生态学报, 23(10): 1951-1962. DOI:10.3321/j.issn:1000-0933.2003.10.001 |
张真. 2003. 马尾松毛虫(Dendrolimus punctatus Walker)种群时空动态及暴发机制研究. 中国科学院动物研究所博士论文, 84-93.
|
张真, 吴东亮, 王淑芬. 1998. 马尾松昆虫群落动态及稳定性研究. 林业科学, 34(1): 65-72. DOI:10.3321/j.issn:1001-7488.1998.01.010 |
张真, 李典谟, 查光济. 2002a. 马尾松毛虫2、3代分化和干旱对种群时间动态的影响. 昆虫学报, 45(4): 471-476. |
张真, 李典谟, 查光济. 2002b. 马尾松毛虫种群动态的时间序列分析及复杂性动态研究. 生态学报, 22(7): 1061-1067. |
张执中, 周章义, 彭建文. 1990. 松毛虫种群动态与环境的关系//陈昌洁. 松毛虫综合管理. 北京: 中国林业出版社, 19-60.
|
邹运鼎, 程扶玖, 查光济. 1990. 松针内含物与马尾松毛虫生存发育关系的研究. 林业科学, 26(2): 142-148. |
Allan D W, John B W. 2002. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Global Change Biology, 8: 1-16. DOI:10.1046/j.1365-2486.2002.00451.x |
Andrewartha H G, Birch L C. 1984. The Ecological Web. Chicago, Illionis: University of Chicago Press.
|
Battisti A, Stastny M I, Buffo E, et al. 2006. A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly global change. Biology, 12(4): 662-671. |
Berryman A A. 1982a. Biological control, thresholds, and pest outbreaks. Environ Entomol, 11: 544-549. DOI:10.1093/ee/11.3.544 |
Berryman A A. 1982b. Mountain pine beetle outbreaks in Rocky Mountain lodgepole pine forests. J For, 80: 410-413. |
Berryman A A. 1987. The theory and classification of outbreaks//Barbosa P, Schultz J. Insect Outbreak. Academic Press, New York.
|
Cappuccino N, Lavertu D, Bergeron Y, et al. 1998. Spruce budworm impact, abundance and parasitism rate in a patchy landscape. Oecologia, 114: 236-242. DOI:10.1007/s004420050441 |
Chitty D. 1971. The natural selection of self-regulatory behavior in animal populations//McLaren I A. Natural Regulation of Animal Populations. New York: Atherton Press, 136-170.
|
Christian J J, Davis D E. 1971. Endocrines, behavior and population//McLaren I A. Natural Regulation of Animal Populations. New York: Atherton Press, 69-98.
|
Denno R F, Gratton C, Peterson M A, et al. 2002. Bottom up forces mediate natural impact in phytophagous community. Ecology, 83(5): 1443-1458. DOI:10.1890/0012-9658(2002)083[1443:BUFMNE]2.0.CO;2 |
Elton C S. 1924. Voles, mice and lemmings. Cramer, Weinheim, U K.
|
Greenbank D O. 1956. The role of climate and dispersal in the initiation of outbreaks of the spruce budworm in New Brunswick Ⅰ. The role of climate. Canadian Journal of Zoology, 34: 453-476. DOI:10.1139/z56-048 |
Hanski I, Saccheri I. 2006. Molecular-level variation affects population growth in a butterfly metapopulation. PLoS Biology, 4(5): e129. DOI:10.1371/journal.pbio.0040129 |
Holling C S. 1965. The functional response of predators to prey density and its role in mimicry and population regulation. Mem Entomol Sco Can, 45: 3-60. |
Hunter A F. 1991. Traits that distinguish outbreaking and nonoutbreaking Macrolepidoptera feeding on northern hardwood trees. Oikos, 60: 275-282. DOI:10.2307/3545068 |
Johnson D M, Bjørnstad O N, Liebhold A M. 2004. Landscape geometry and travelling waves in the larch budmoth. Ecology Letters, 7(10): 967-974. DOI:10.1111/ele.2004.7.issue-10 |
Lotka A J. 1925. Element of Physical Biology. Baltimore, Marylan: Williams & Wilkins.
|
Mattson W J, Haack R A. 1987. The role of drought stress in provoking outbreaks of phytopherogous insect//Barbosa P, Schultz J C. Insect Outbreaks. Academic Press, New York.
|
Morris R E. 1963. The dynamics of epidemic spruce budworm populations. Mem Entomol Sco Can, 31: 1-332. |
Nicholson A J, Bailey V A. 1935. The balance of animal populations. Proc Zool Soc London Pt. 1: 551-598.
|
Nylin S. 2001. Life history perspectives on pest insects: What's the use?. Austral Ecology, 26(5): 507-517. DOI:10.1046/j.1442-9993.2001.01134.x |
Radeloff V C, Madenoff D J, Boyce M S. 2000. The changing relation of landscape patterns and jack pine budworm populations during an outbreak. Oikos, 90: 417-430. DOI:10.1034/j.1600-0706.2000.900301.x |
Redfearn A, Pimm S L. 1987. Insect Outbreaks and Community Structure//Barbosa P, Schultz J. Insect Outbreaks Academic Press, New York.
|
Rhoades D F. 1985. Offensive-defensive interactions between herbivores and plants: Their relevance in herbivore population dynamics and ecological theory. American Naturalist, 125: 205-238. DOI:10.1086/284338 |
Southwood T R E, Comins H N. 1976. A synoptic population model. Journal of Animal Ecology, 65: 949-965. |
Spitzer K, Rejmanek M, Soldan T. 1984. The fecundity and long-term variability in abundance of noctuid moths (Lepidoptera: Noctuidae). Oecologia, 62: 91-93. DOI:10.1007/BF00377379 |
Takahasi F. 1964. Rproduction curve with two equilibrium points: A consideration of the fluctuation of insect population. Res Popul Ecol, 6: 28-36. DOI:10.1007/BF02524942 |
Watt K E F. 1964. Comments on fluctuations of animal populations and measures of community stability. Can Entomol, 96: 1434-1442. DOI:10.4039/Ent961434-11 |
Watt W B, Wheat C W, Meyer E H, et al. 2003. Adaptation at specific loci. Ⅶ. Natural selection, dispersal and the diversity of molecular-functional variation patterns among butterfly species complexes (Colias: Lepidoptera, Pieridae). Mol Ecol, 12: 1265-1275. DOI:10.1046/j.1365-294X.2003.01804.x |
Wellington W G. 1960. Qualitative changes in natural populations during changes in abundance. Canadian Journal of Zoology, 38: 289-314. DOI:10.1139/z60-036 |
Wheat C W, Watt W B, Pollock D D, et al. 2006. From DNA to fitness differences: Sequences and structures of adaptive variants of Colias phosphoglucose isomerase (PGI). Mol Biol Evol, 23: 499-512. DOI:10.1093/molbev/msj062 |
White T C R. 1978. The importance of a relative shortage of food in animal ecology. Oecologia, 33: 71-86. DOI:10.1007/BF00376997 |
Zhang Z, Li D M. 1999. A possible relationship between outbreaks of oriental migratory locust (Locusta migratoria manilensis Meyen) in China. Ecological Research, 14: 267-270. DOI:10.1046/j.1440-1703.1999.t01-1-143305.x |