 2008, Vol. 44
2008, Vol. 44文章信息
- 樊敏, 马履一, 王瑞辉.
- Fan Min, Ma Lüyi, Wang Ruihui
- 刺槐春夏季树干液流变化规律
- Variation of Stem Sap Flow of Robinia pseudoacacia in Spring and Summer
- 林业科学, 2008, 44(1): 41-45.
- Scientia Silvae Sinicae, 2008, 44(1): 41-45.
-
文章历史
- 收稿日期:2006-11-03
-
作者相关文章


刺槐(Robinia pseudoacacia)生长速度快、耐干旱、耐瘠薄,现已成为干旱半干旱地区的水土保持树种和主要造林树种(茹桃勤等,2005;单长卷,2005),同时也是城市绿化的优良树种,在植被恢复和生态建设中发挥着重要的作用。
树木在发挥巨大生态效益的同时,维持自身生长发育需要消耗大量水分(王华田,2003),而水资源缺乏已经成为当今全球日益突出的问题,因此,树木的水分传输规律特别是整株树木的耗水量受到国内外树木水分生理学家、生态学家、林学家和园林学家的共同关注(马履一等,2003;苏建平等,2004;Tatarinov et al., 2005)。由于树干边材液流速率及液流量制约着整株树木的蒸腾量,因而对树干液流进行标记并测定其流动速率就可以简捷地确定树冠蒸腾耗水量(李海涛等,1998;高岩等,2001)。热扩散技术是在基本保持树木自然生活状态不变的条件下,将微型热电偶传感器插入边材木质部中,根据电热转换和能量平衡原理测定树干边材液流的运移速率(Linton et al., 1998),进而利用被测部位的边材横断面积求得单木整株液流量(Granier et al., 1994;Wullschleger et al., 1998;马履一等,2002)。因为这种方法简便易行、测量精度高,所以国内外越来越多的学者投入到边材液流的研究中(王瑞辉等,2006),但以往的研究通常选择栽植在山地或丘陵等宜林地上的树木边材液流进行阶段性观测,尚未见对栽植在城市中建筑渣土上的树木边材液流进行长期连续观测,亦未见对栽植在这一环境条件下的刺槐树干液流长期观测研究的报道。本文采用热扩散式茎流计对生长在城市中建筑渣土上的刺槐树干液流进行长期观测,旨在揭示城市环境下刺槐树干液流传输的变化规律,以期为北方城市园林绿化低耗水树种的选择、结构配置和绿地的合理灌溉提供理论依据。
1 试验地概况及研究方法 1.1 试验地概况试验地设在北京林业大学主楼前的绿地内。北京林业大学位于北京市西北部,海拔75 m,属暖温带半湿润季风型大陆性气候,年平均气温11~15 ℃,极端最高气温达40 ℃,极端最低气温达-15 ℃,平均年降水量为600 mm左右,年蒸发量为1 835.8 mm。主楼前绿地内的表层土为轻壤土,厚35 cm,表土以下为建筑渣土; 地上树木疏林状种植,林下有草地早熟禾(Poa pratensis)覆盖。
1.2 研究方法按照树干通直、生长良好、不偏心、不偏冠的要求在试验地内选择4株不同大小的刺槐样株作为研究对象(表 1)。样木1为林分平均木,样木2~4用于径阶试验。
|
|
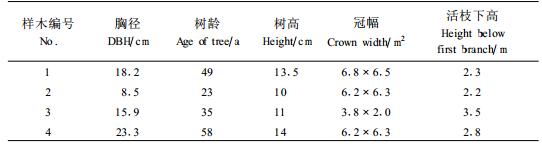
于2006年4月1日—8月31日共5个月对各样木的边材液流进行连续同步测定。为了避免因树干不同方位液流的差异而导致研究结果产生严重偏差,所以将TDP探针统一安装在每株样木树干1.3 m处的南向。首先在探针安装处刮去样木的粗皮,然后采用特定规格的钻头,依据仪器自带模具沿树干纵向垂直钻取直径1.5 mm的孔洞,插入TDP-30探针。将所有探针的另一端与数采器(Data Logger)连接,同时,在样地内安装自动气象站,同样与数采器连接,实现树干液流和气象因子数据的同步自动采集,数据采集的间隔期为10 min。利用生长锥测量各样木的边材面积,应用Dynamax公司提供的软件和SPSS13.0统计软件对所有数据进行分析。
2 结果与分析 2.1 边材液流速率的日变化及季节变化动态由于在4—8月中,刺槐各日液流速率的日变化图形非常密集,很难直观地对其日变化及季节变化动态进行具体分析与比较,所以从春季(5月)和夏季(7月)中分别选择3个连续的晴天,用所选各日样木1的树干液流速率测定结果绘制图 1。从图 1中可以看到,刺槐边材液流的日变化为典型的宽峰形曲线,每日从6:50左右启动,13:00左右达到峰值,液流达到峰值后仍有较小幅度的“波动",形成多个小峰组成的“高峰平台",这与孙鹏森(2000)、王华田(2002)对刺槐树干液流的研究结果一致。19:30左右开始迅速下降,直至来日液流启动之前降至低谷,没有明显的液流停止界限,夜间仍有较高的液流存在,这主要是由根压引起的。根压使水分以主动吸收的方式进入树木体内, 补充白天蒸腾丢失的大量水分, 恢复树木体内的水分平衡(高岩等,2001)。

|
图 1 刺槐树干液流速率的日变化及季节变化 Figure 1 Diurnal and seasonal fluctuation of sap flow velocity of R. pseudoacacia |
表 2列出了所选各日几个表达曲线特征的指标,从启动时间来看,7月启动较早,为6:40,5月启动较晚,为6:50,即夏季(7月)液流启动的时间要早于春季(5月)。液流启动的这种变化规律与太阳辐射开始增强的规律相吻合。所选观测日太阳辐射开始增强的时间分别是:5月13日4:50、5月14日4:50、5月15日4:40、7月14日4:30、7月15日4:30、7月16日4:30,显然,7月(夏季)太阳辐射开始增强的时间早于5月(春季),与液流启动变化规律一致。从生理学的角度来看,清晨,随着太阳辐射的逐渐增强,空气温度逐渐升高,诱导刺槐叶片气孔张开,光合与蒸腾逐渐增强,产生的蒸腾拉力带动液流启动(武维华,2003),夏季太阳较春季出来早,所以液流启动早。
|
|
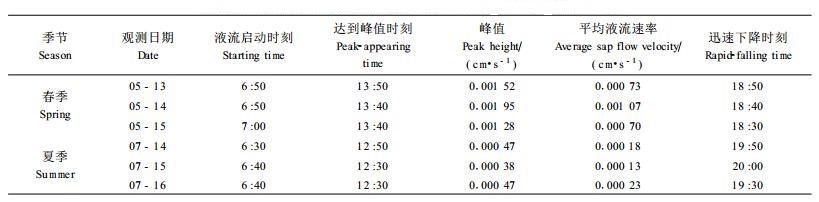
从达到峰值的时间来看,7月较早,12:40左右出现,5月较晚,13:50左右出现,即夏季(7月)早于春季(5月)1 h左右,与液流启动变化规律一致; 从峰值和日平均液流速率来看,7月较小,连续3个观测日的峰值平均为0.000 44 cm·s-1,日平均液流速率为0.000 18 cm·s-1,5月较大,峰值平均为0.001 58 cm·s-1,日平均液流速率为0.000 83 cm·s-1,显然,夏季(7月)的峰值和日平均液流速率均小于春季(5月),分别为春季的27.8%和21.7%;从液流开始迅速下降的时间来看,7月较晚,为19:50左右,5月较早,为18:40左右,总趋势是夏季(7月)晚于春季(5月)1 h左右。
综上所述,夏季的液流启动时间和达到峰值的时间均早于春季,而迅速下降的时间又晚于春季,显然,夏季液流高峰维持的时间长于春季,即夏季液流速率的曲线峰形明显宽于春季,但是夏季的峰值和日平均液流速率却小于春季。
2.2 边材液流通量的月变化及季节变化动态将2006年4—8月连续观测到的样木1的液流速率按月统计平均数,计算平均液流通量,绘制图 2。从图 2可以看到,刺槐的平均液流通量4月最大,为4.075 cm3·h-1cm-2,6月最小,为1.764 cm3·h-1cm-2。虽然4月的平均液流通量最大,但是月度总耗水量不一定最大,因为刺槐4月的实际耗水时间仅为5 d,在4月26日之前的液流很小,仪器检测不到,直到26日以后才急剧上升。如果按季节比较,4、5月为春季,6、7、8月为夏季,则刺槐春、夏季的平均液流通量分别为3.042 cm3·h-1cm-2、2.403 cm3·h-1cm-2,显然春季>夏季。

|
图 2 刺槐平均液流通量的月变化及季节变化 Figure 2 Monthly and seasonal fluctuation of average sap flux of R. pseudoacacia |
春季液流通量大的原因是多方面的、综合性的。一是与刺槐的生长发育有关,春季是刺槐抽梢、展叶和开花的季节,对水分的需求很大; 二是与北京地区春季干旱多风、雨天较少的特殊气候特征有关。北京春季风速比夏季大,与液流同步监测的气象数据显示,春、夏季平均风速分别为0.468 m·s-1和0.338 m·s-1; 春季空气湿度比夏季低,平均为48.27%, 夏季平均为66.85%;春季雨天比夏季少,多为晴天和多云。风速的增加使紧贴叶子表面的空气层变薄,水汽扩散阻力减小,蒸腾加快,空气湿度的降低使大气水势减小,大气与叶片气孔下腔间的水气压梯度加大,蒸腾耗水的速率加快,而晴天的蒸腾耗水速率又显著大于阴雨天,所以春季的液流通量较大(徐军亮,2006;王瑞辉,2006)。
2.3 不同直径树干边材液流的变化动态从样木1~4的6月份观测数据中分别选择3个连续的晴天,用所选各日的树干液流速率绘制图 3,并将其相应的几个表达曲线特征的指标分别列入表 3,分别求算样木1~4在所选各日中的耗水量绘制图 4。

|
图 3 刺槐不同直径树干液流速率的日变化 Figure 3 Diurnal fluctuation of sap flow velocity of R. pseudoacacia in diametric classes |
|
|


|
图 4 刺槐日耗水量随直径的变化规律 Figure 4 Diurnal fluctuation of water consumption in diametric classes |
从图 3和表 3中可以看出,不同直径刺槐的边材液流速率在日周期内的启动时间是基本一致的,均在6:30左右,但达到峰值的时间、峰值的大小、日平均液流速率和迅速下降的时间差异很大。直径8.5 m单株达到峰值时间明显晚于直径15.9、18.2、23.3 cm的单株4~5 h,而直径15.9 cm单株的达到峰值时间又明显早于18.2、23.3 cm的单株1 h,直径18.2 cm和23.3 cm的单株达到峰值时间基本一致,均在9:20左右; 直径8.5 cm的单株,峰值和日平均液流速率显著大于其余3株被测单株,直径15.9、18.2、23.3 cm单株的峰值和日平均液流速率基本相同; 直径15.9 cm单株的迅速下降时间显著晚于其余3株被测单株2~2.5 h,直径8.5、18.2、23.3 cm的单株,迅速下降的时间基本一致,均在17:50左右。可见,刺槐边材液流速率的变化与树干直径关系不大。值得注意的是,直径8.5 cm刺槐的边材液流速率曲线峰形较窄,其余3株直径较大的刺槐边材液流速率曲线峰形较宽,这可能是由于树体水容在树冠蒸腾过程中的调节作用造成的。直径较小的刺槐,树体水容小,可供蒸腾的水少,故峰形较窄; 反之,直径较大的刺槐,树体水容大,可供蒸腾的水多,故峰形较宽。
从图 4可以看出,刺槐单株的日耗水量是随直径的增大而增加。直径23.3 cm的单株耗水量最大,日平均为9.751 kg; 直径18.2 cm和直径15.9 cm的2株刺槐耗水量居中,日平均分别为7.874 kg和6.744 kg,分别为直径23.3 cm单株耗水量的80.8%和69.2%;直径8.5 cm的单株耗水量最小,日平均仅4.149 kg,为直径23.3 cm单株耗水量的42.5%,直径18.2 cm单株耗水量的52.7%,直径15.9 cm单株耗水量的61.5%。用SPSS13.0软件进行相关因子分析,结果表明,刺槐单木日耗水量与树干直径和边材面积的相关性达极显著水平,相关系数分别为0.983和0.999,与王华田(2002)在侧柏(Platycladus orientalis)上的试验结果和王瑞辉(2006)在元宝枫(Acer truncatum)上的试验结果相同。
3 结论刺槐春夏季边材液流的日变化呈宽峰形曲线,每日6:50左右启动,13:00左右达到峰值,19:30左右开始迅速下降,没有明显的液流停止界限,夜间仍有较高的液流存在。另外,研究发现,刺槐夏季液流每天启动的时间早于春季10 min左右,达到峰值的时间早于春季1 h左右,峰值和日平均液流速率小于春季,迅速下降的时间晚于春季1 h左右。
刺槐在4—8月期间,各月的平均液流通量如果按月比较则4月最大,为4.075 cm3·h-1cm-2,6月最小,为1.764 cm3·h-1cm-2; 如果按季节比较,则春季的平均液流通量为3.042 cm3·h-1cm-2,显然大于夏季的2.341 cm3·h-1cm-2。刺槐边材液流速率和液流通量的季节变化说明刺槐春季蒸腾量大,是水分管理的关键时期。了解和掌握了刺槐的日耗水规律和季节耗水规律,就可以结合同步监测的环境因子构建液流通量与环境因子的关系模型,进而根据北京市气候和土壤水分状况的季节和年周期变化规律,预测和计算刺槐单株日、月和季节的蒸腾耗水量和需水量,提出精准的灌溉方案,制定合理的灌溉制度。
刺槐不同直径边材液流速率的变化与树干直径关系不大,直径小的边材液流速率曲线峰形较窄,直径大的边材液流速率曲线峰形较宽; 日周期单木耗水量随树干直径的增大而增加,与树干直径和边材面积相关显著,相关系数分别为0.983和0.999,这为实现刺槐从单木到群体的尺度扩展提供了条件。在研究林木群体耗水量时,只要找出群体内单木耗水量与边材面积随直径的分布规律,就可以估算群体的现实耗水量,从而实现单木到群体的尺度扩展(马李一等,2001;Cermak et al., 2004)。
高岩, 张汝民, 刘静. 2001. 应用热脉冲技术对小美旱杨树干液流的研究. 西北植物学报, 21(4): 644-649. DOI:10.3321/j.issn:1000-4025.2001.04.007 |
李海涛, 陈灵芝. 1998. 应用热脉冲技术对棘皮桦和五角枫树干液流的研究. 北京林业大学学报, 20(1): 1-6. DOI:10.3321/j.issn:1000-1522.1998.01.001 |
马李一, 孙鹏森, 马履一. 2001. 油松、刺槐单木与林分水平耗水量的尺度转换. 北京林业大学学报, 23(4): 1-5. |
马履一, 王华田, 林平. 2003. 北京地区几个树种耗水性比较的研究. 北京林业大学学报, 25(2): 1-7. |
马履一, 王华田. 2002. 油松边材液流时空变化及其影响因子的研究. 北京林业大学学报, 23(4): 23-27. DOI:10.3969/j.issn.1671-6116.2002.04.004 |
茹桃勤, 李吉跃, 孔令省, 等. 2005. 刺槐耗水研究进展. 水土保持研究, 12(2): 135-140. DOI:10.3969/j.issn.1005-3409.2005.02.042 |
单长卷. 2005. 土壤干旱对刺槐幼苗水分生理的影响. 安徽农业科学, 33(10): 1852-1853. DOI:10.3969/j.issn.0517-6611.2005.10.039 |
苏建平, 康博文. 2004. 我国树木蒸腾耗水研究进展. 水土保持研究, 11(2): 177-186. DOI:10.3969/j.issn.1005-3409.2004.02.057 |
孙鹏森.2000.京北水源保护林树种不同尺度耗水特性及林分配置的研究.北京林业大学博士学位论文.
|
王华田.2002.北京市水源保护林区主要树种耗水性研究.北京林业大学博士学位论文.
|
王华田. 2003. 林木耗水性研究述评. 世界林业研究, 16(2): 23-27. DOI:10.3969/j.issn.1001-4241.2003.02.005 |
王瑞辉, 马履一, 奚如春. 2006. 元宝枫生长旺季树干液流动态及影响因素. 生态学杂志, 25(3): 231-237. DOI:10.3321/j.issn:1000-4890.2006.03.001 |
王瑞辉.2006.北京主要园林树种耗水性及节水灌溉制度研究.北京林业大学博士学位论文.
|
武维华. 2003. 植物生理学. 北京: 科学出版社, 47-66.
|
徐军亮.2006.京西山区油松、侧柏单木耗水环境影响因子评价与模拟.北京林业大学博士学位论文.
|
Cermak J, Kucera J, Nadezhdina N. 2004. Sap flow measurements with some thermodynamic methods, flow integration within trees and scaling up from sample trees to entire forest stands. Trees Structure and Function, 16: 529-546. |
Granier A, Anfodillo T, Sabatti M, et al. 1994. Axial and radial water flow in the trunks of oak trees:a quantitative and qualitative analysis. Tree Physiology, 14(12): 1383-1396. DOI:10.1093/treephys/14.12.1383 |
Linton M J, Sperry J S, Williams D G. 1998. Limits to water transport in Juniperus osteosperma and Pinus edulis: implications for drought tolerance and regulation of transpiration. Functional Ecology, 12(6): 906-911. DOI:10.1046/j.1365-2435.1998.00275.x |
Tatarinov F A, Kucera J, Cienciala E. 2005. The analysis of physical background of tree sap flow measurement based on thermal methods. Measurement Science and Technology, 16: 1157-1169. DOI:10.1088/0957-0233/16/5/016 |
Wullschleger S D, Meinzer F C, Vertessy R A. 1998. A review of whole-plant water use studies in trees. Tree Physiol, 18(8/9): 499-512. |