


在生态学诞生初期,英国科学促进会时任主席于1893年把生态学描述为与生理学和形态学并列的3个生物学分支学科之一,并且认为是“最有吸引力的”[1]。
2010,《Science》亚太主编 Richard Stone撰写报告,指出生物学是当代自然科学中比重最大的学科,而生态学又是在全部生命科学论文中数量最多的研究领域之一[2]。一般而言,生态学被普遍地看作是后现代性科学而被认为不同于以近代物理学为代表的经典学科[3]。这种学术观点在高德史密斯(E.Goldsmith,1992)的《The Way an Ecological World-view》一书中表现得淋漓尽致,鉴于高德史密斯在学界和社会各界的直接与间接的重大影响,这种学术观点的影响广泛与深远可见一斑。高德史密斯引用贝塔郎菲(Ludwig Von Bertalanffy)的话,认为存在“介于物理学和生态学两者的基本不同”,并认为生态学中存在着不同于物理学和化学的概念,“迄今物理学与化学定律仍旧被关注……但没有这些和相似的概念,就没有医学和真正的生物学科学。”在《The Way an Ecological World-view》第11章引用维斯(Paul Weiss)的话而主张“‘没有理由将自然降格’去迎合物理学家的不足”[4]。
但生态学的地位和自身存在问题的严重性并不一致。继爱因斯坦以后,物理学界仍旧期待着统一理论(此处作等同于“一般理论”的理解)的出现并持续探索,而生态学界对于是否可能存在一般理论还存在着质疑。2009年,Roughgarden在《Biology and Philosophy》上发表论文,对群落生态学存在一般理论(general theory)的观点提出了质疑。他感慨:“我欣赏建构作为一般理论的理论群落生态学的呼吁。如果谨慎的睁开眼睛寻找一般理论,我们能避免(错误)投入一般理论的危险。所以,今天群落生态学有一般理论吗?没有。可能有吗?是的。应该有吗?我不知道。”[5]事实上,不仅是群落生态学是否具有一般理论的可能存在争鸣,即使对于整个生态学而言,是否存在一般理论都没有达成一致[[6]。即使是达尔文的演化说和自然选择法则,也被 Popper(1959,1972,1983)批判了[7]。
当前,绝大部分生态学家都回避了达尔文式的、对于一般理论的探寻,转而研究具体的生态过程,将生态圈切割成了纵横交错的巨大区组(plot)群并分而击之,诞生了植物生态学、动物生态学、微生物生态学、海洋生态学、森林生态学等。生态学从大写的ECOLOGY变成小写的ecologies。
作为科学的一部分,生态学也注重实验。对此,国内一些生态学家和生态学哲学家从实验方法角度探寻了导致影响生态学一般理论出现的原因,不过人数与成果零星,且研究或较不系统或仅针对诸如“伪重复”某一个具体原因[8~11]。相较于国内,《Nature》与《Science》杂志均已以社论或调研报告的形式提出生态学研究存在实验结果的重复性差的问题,这暗示了生态学研究需审慎提出包括一般理论在内的各种假说。但文献检索表明,这个问题并没有得到生态学家的足够重视并给予针对性的研究。
2 生态学假说的判决性实验生态学到底有没有可能像物理学一样努力找到一般理论?首先应该明确理论应该满足什么条件。1905年,爱因斯坦提出了广义相对论,因为没有被实验验证,最初只是物理学假说,一直到1919年,艾丁顿勋爵在非洲对于日全食的观察实验得到的数据与广义相对论的预言高度吻合,物理学界才把广义相对论从未经严格验证的假说升格为作为具有普遍性的理论。换言之,只有经过严格验证的假说才能成为理论。艾丁顿勋爵的观察实验验证的思路很简洁:以爱因斯坦的广义相对论假说作为H0,以不符合广义相对论假说的所有其他可能的结果作为H1,如果实验结果支持H0,则广义相对论可以成为理论,如果实验结果支持H1,则广义相对论不仅不可以成为理论,甚至连假说的资格也被取消。这种非此即彼的实验验证方法称之为判决性实验。
由此,参照物理学的思路,探寻生态学理论的有效方法之一是对于各种假说进行判决性实验验证。通过实验对假说进行判决性验证的方法受到了很多研究者的认可[12~14]。2000年,Scheiner和Gurevitc在《生态学实验设计与分析》首页便提出:“生态学日益发展成为一门实验科学……生态学家们越来越多地利用实验来验证自然的理论(Hairston,1989;Resetarits和Bernardo,1998)。”[15]实验方法包括实验室实验和野外实验两种类型。哪一种类型适合生态学的研究特点并能胜任假说的判决性实验,需要逐一考察两者。
3 生态学假说的实验室验证实验室验证在物理学取得成功在于2个原因。
首先是因为它的理论体系建立在机械论的自然观基础之上,研究对象都是自然界中的非生命事物,比如: 中国大众最为熟知的可能是已经被证明是杜撰的伽利略在比萨斜塔的自由落体实验——在这个实验中,所有对象都是机械性的,没有生命。
其次,物理学实验室验证采取了单因子假设,即仅仅对于影响受试对象的一个影响因子的初始值进行操控(manipulate),观察该影响因子对于受试对象的作用,验证相关的物理学假说。因为经典物理学对象的机械性,其物理特性不仅不会因为在实验室中而产生与在自然环境中不同的变化,而且,在同样的条件下,任何实验室的任何一次实验都应该呈现一致的结果,这就是实验的可复制性——在中国的实验室应该与在美国的实验室有着一样的结果。换言之,物理学实验室其实就是一个“微宇宙”(micro-cosmos)——其中发生的一切和真实的物理世界应该完全一致。艾丁顿勋爵的观察实验其实就是采取了单因子假设的思路,即只有广义相对论这一个影响因子导致了大尺度的空间弯曲现象的产生,而没有假设还存在其他可能的影响因子。
生态学家正在努力地进行着各种假说的实验室验证,但进程并不顺利。
不同于物理学,生态学的研究对象往往是活的生物,影响受试对象的影响因子有时也是活的生物,这就使得实验室实验的操控非常困难。更重要的是,生态学研究的是“生物及与环境之间的关系”[16],而生态过程又始终处于演化之中,这决定了实验的受试对象的响应模式可能是多样的,既可能是基于单因子假设(如猎物-捕食者模型(Lotka-volterra)),也可能是基于多因子假设(如植物的生态演替(ecological succession));影响受试对象的影响因子这种关系可能是网状的(web),如食物链;也可能是层次等级的(hierarchy),如全球水循环。简而言之,生物学研究对象的影响因子及关系比物理学复杂得多,这种复杂性可能导致实验的失败或不可复制,甚至于,研究者本身没意识到导致实验成功的主导因子(dominantfactor)及关系——比如,有可能把实验中在时间序列上呈现的先后关系错会成了逻辑上的因果关系,把非原假设当作原假设(null hypothesis) 处理了。
2007年,Svensson等指出,在实验室验证的实验方案,设计时没有采用单因子假设选择干扰项(agent),导致不同物种和功能群体对于实验处理的响应不同,进而无法进行假说的判决性验证[[17]。2000年,Radford和Cousens对Rejmanek于1996年提出的“植物入侵的一般理论”生态学假说进行了判决性实验,他们采用了单因子假设的思路,比较了入侵的S.madagascariensis和没有入侵的S.lautus的情况。但他们不仅通过实验室验证否定了假说,并进一步否定了通过假说的判决性实验验证探寻生态学理论的方法,他们认为“期待发展出对全部生境全部物种的预测系统是不合理的”结论[18]。他们否认判决性实验方法的主要理由是生态学研究对象与物理学研究对象的差异。
2005年,Ernest研究了生态学与物理学的差异,认为物理学的实验室验证可以单独去除所有使得数据辨识度降低的外部因素,但在生态学实验中不太可能这样操作,这和Chatfield于1995年提出导致统计误差的3个原因之一有关——在组分(composite)和数量都已知的实验对象中出现了假说无法解释的随机变量(即影响因子),这种随机变量(比如物种组成、土壤湿度和海拔等)对于生物和非生物影响是不同的,进一步的,不同的实验室实验是在特定的生物或非生物地点进行操作,必然产生更加大量的误差[19]。也就是说,出现了有待实验验证的影响因子以外的其他影响因子(多影响因子)。
所以,实验室方法不可能还原整个自然界所有的影响因子与关系(不然,这个实验室就必须在物理空间上具有和生态圈一样的尺度(scale)与范围(domain)),肯定不适合作为生态学假说的实验验证方法。
4 生态学假说的野外实验验证相对于实验室实验,野外实验在生态学中日益流行。2013年,林祥磊将生态学各种具体实验方法进行分类研究后,认为野外实验“可以称为原地实验,是指在野外条件下采取某些措施获得某些生态因素的变化对生态学系统及其他因素的影响的实验。……野外实验是在更接近实验对象存在和生长的环境条件下进行的,因此,检测到的交互作用显然也是在自然界中发生的,对于研究大的、存活时间长的,或行为复杂的生物,野外实验是唯一可行的方法,在实验室的限制下不可能对它们进行验证”[[20]。
但野外实验,一方面,同样存在关于实验控制和单/多因子的争鸣。
4.1 关于实验控制的争鸣与处理2000年,Scheiner和 Gurevitch认为野外实验操作时有成本高昂、难以重复和数据“噪音”(noise) 三大困难。导致上述困难的主要原因是生态学研究对象处于演化中,实验设计、操作与记录等都很难对于生态现象的演化作出响应——“自然界的变化经常掩盖了我们试图检测的处理结果”[15]。Jaeschke等 2014的研究表明“野外研究正在强劲增长”,但因为研究对象始终处于演化中的整个自然界,对于野外实验的 方法适用也提出了质疑,认为它无法解决由生境的多样性导致的生态过程的多样性与试图对所有类似的生态过程进行解释的矛盾[19]。这些文献集中反映了对于生态学野外实验中控制实验对象(活的生物)的困难。
若野外实验的条件完全符合实验所要验证的生态学假说且没有额外的干扰项,则野外实验一定可以对于假说进行判决性实验。此时,野外实验本质上和实验室的受控实验没有区别了,无非是实验环境的物理空间的范围差异,并非尺度差异。生态过程的复杂性使得对其的了解非常少,且生态过程在不断演化,其中的影响因子以及相互关系难以发现和确定,所以无法严格地认为被选择进行野外实验的实验地点就恰好符合实验的要求。但这也说明了野外实验具有自然性,是自然的一部分,野外实验本身就可以近似地看做是生 态过程的一部分(只是这个生态过程包括了人类这个影响因子)。所以,野外实验还不完美,但却是已知实验方法中最适合进行假说的判决性实验的方法。
4.2 关于单/多因子假设的争鸣与处理2005年Holyoak和Lawler反驳了Srivastava等于2004年的研究——他们在实验室用自养微生物进行微宇宙实验,验证了关于种群竞争的逻辑斯蒂(logistic)方程和捕食者-猎物(Lotka-Volterra)模型,发现对于采用单因子假设的、代际更替周期短的种群动力学研究不适宜采用环境复杂且周期长的野外实验[21]。2008年,Griffen和Drake对种群灭绝的文献进行分析研究,涉及了Web of Science自1956年到2007年7月的所有文献和部分早于1956年发表的被引用文献,从1016篇文献中选取了53篇进行研究,发现现有理论绝大部分是基于单因子假设的,同时以亚种间和物种间的交互作用为代表的多因子假设也有很大影响[22]。换言之,单因子假设与多因子假设目前都存在于生态学研究中。在这种情况下,如果野外实验同时适用单因子假设和多因子假设的实验方法,就可以避免陷入这方面无尽讨论,让作为方法的生态学假说判决性实验验证成为可能。
1)基于单因子假设的野外实验验证的思想实验。
以生态学研究中典型的群落(community)研究为例。波动(pulse)和压力(press)是生态过程中常见的两种扰动(perturbation)。波动扰动是群落演替过程中由外部产生的较为剧烈但有规律的物理扰动,比如:温度和光照等在自然界中往往存在周期性变化的因子对于植物生长的影响。压力扰动是群落内的种群密度改变后对于整个群落的影响,比如:在捕食者-猎物模型中,若捕食者过渡捕猎导致猎物减少,最后的结果是捕食者数量也会因食物短缺减少。若生态过程是单因子决定的,则两种扰动只有一种才会决定群落的演化。野外实验可对有n个种的群落通过单独展开两种扰动改变种的某一亚种组(subset)密度的实验来明确哪种扰动才是影响群落的主导因子。
波动扰动实验首先测定n个种的亚种(包括被扰动种)的初始增长率。在密度变化时,其他种会反复调节其密度。这种调节结束以后,群落会达到新的平衡。在这一平衡时,可以测定多个种的密度净变化。野外实验一个长时间未解决的难题是确定实验对象在分类学上的地位。在波动扰动实验中,这个问题可以通过把所有物种看作是亚种组S,通过做S次实验,求出所有的结果。这个处理方法符合Platt(1964)的实验4步骤要求[23]。压力实验通过多次迁出被扰动种的个体使其密度保持在预先规定的水平,被扰动种的密度也维持在平衡水平。在资源充沛前提下,各个物种相互可以有较高的平衡密度,种间基本不存在竞争。不过,生态过程是普遍存在关系的(生态学的基本定义),当迁入新的物种个体后,平衡密度就被打破。这个处理方法也符合Platt于1964年提出的实验4步骤要求[23]。
Bender等1984年的成果表明,如果分别进行波动扰动和压力扰动实验,会发现在压力扰动实验中,当一个物种或另一个物种被迁出或实验保持在某一低密度时,2个激烈竞争的物种可能不表现出密度的净增加。这是由于种间潜在的反复缓冲作用,尤其在资源动态变化非常缓慢时[24]。他们的成果证明波动是决定群落中种群密度的主导因子——这说明野外实验可以作为基于单因子假设的实验验证方法。
2)基于多单因子假设野外实验验证的实验设计。
景观(landscape)生态学是20世纪90年代兴起的生态学分支,核心概念是“补缀”(patch),它是“不同于它所处的自然环境的、具有同质性的、离散空间模式中有界的、联系的不连续体”[16,25]。补缀的定义决定了在补缀任何一处进行的实验条件均保持了一致性。若生态过程是多因子决定的,只需对同一补缀的多个地点分别展开多因子的野外实验即可确定哪些(而不是哪一个)因子是主导因子(即A地点验证a因子B地点验证b因子)。换言之,具有相同性质和特点的补缀在更大的空间中构成了景观,多因子可分别在各个补缀中同时展开实验,最终找到决定生态过程的多因子并明确关系。所以,景观适合作为多因子模型假说的实验验证对象。
具体而言,对景观进行生态学假说的野外实验要求不同的实验单元接受2种以上的不同处理,并且对实验单元的处理分配是随机的。由于采用对照、重复、实验操作的随机化和分散安排实验单元等手段来控制偏差和随机误差,因此结论较为可靠。实验设计要求选择的景观大小适合研究对象的时空尺度,不同实验单元接受不同的、随机或分散安排的处理,并且每一种处理的实验单元有足够重复,还要安排时、空对照来排除外来因素的干扰,且为确保观测到实验的滞后效应,处理后的取样时间必须足够长。这样可确保实验单元 的均质性和一致性,实验结果的差异才可直接归因于处理的效果[26]。
由此,野外实验也可以作为基于多因子假设的实验验证方法。
5 生态学野外实验的新操作规范野外实验和实验室实验都是特定的实验方法,应该具有相应的操作规范。受益于物理学的奠基,实验室实验的操作规范非常系统、成熟,但野外实验的操作规范就不尽然,在生态学应用中更是引起了争鸣。
2010年,Williams等报告了对解释外来物种在新生境中高密度生长机制的两种假说的野外实验验证的结果,他们承认因为区组(block)过少(3个),研究结论不适合作为假说的验证结果[27]。他们的研究反映了现有野外实验操作规范不能胜任生态学假说的判决性实验验证的现状。Jaeschke等在2014年总结性地概括野外实验验证失败的原因在于“不能包括所有的多样性和空间”[19]。换言之,因为生态圈涉及从大气圈到地表以下1km处的所有自然界事物,即使是在野外进行实验,也不确保A地的构成要素及交互作用与其他地区相同,形成的实验结果就可能与其他地区的不尽相同,自然不能形成放之四海皆准的结论,也就不能验证生态学的假说。这要求生态学假说的野外实验验证具有完善的操作规范。
进一步的,操作规范不仅是实验控制的具体技术手段,也是单因子/多因子实验有效的前提之一。
以单因子假设的波动扰动实验为例,群落生态学家不可能测定出群落中每一个潜在相互作用种的密度, 而且容易把某些种和另一些种集合成大类(composite category),这可能把间接作用和交互作用混淆。很难规定怎样才能寻找出种间相互作用激烈的一组种群。在压力实验中,因为一个种对另一个种的重要程度可能不取决于其体躯大小、分类地位或生物量,所以,即使是微小的寄生物都可能从根本上改变测定的选择性寄主之间的交互作用。以上情况,都可能导致实验的失败,使得野外实验无法进行有效的假说验证[23]。对此,只有采取统一的分类标准,才能避免这些问题的出现。
以多因子假设的景观生态为例,实验验证的成功将受到以下方面的制约:实验单元内部和彼此之间的空间异质性难以保证真正的重复;野外很难控制多个独立的变量;研究对象的大尺度可能给实验操作带来难以克服的困难;现实操作易受经费、人力、设备不足,实验对象所有权限不同等管理等因素的限制[26]。其中,空间异质性恰巧有助于验证假说是否正确并且是否一般。换言之,空间异质性使得假说的可证伪性进一步提高,获得证实的假说具有更大的解释力。野外环境和大尺度(scale)对于实验造成的困难需要采取严谨和一致的操作才可能尽可能排除无关的干扰。这些都和生态学假说的野外实验验证的操作规范有关。
野外实验需要发展出不同于实验室实验的完整、独立的规范,才可能在现实中完成对生态学假说的严格验证。鉴于生态学研究对象存在于整个生态圈,假说的野外实验验证也需要在多个尺度和不同范围内进行实验,实验结果才有望确保结论具有一般性。仅就逻辑而言,应该在整个生态圈的所有类型的生态位和水平上进行同样的野外实验操作,才能确保被验证的假说被完全证实。显然,这样的操作规范与从实验室操作规范借鉴而来的传统野外实验操作规范应有极大的差异。
2013年,Fraser等在研究全球生态时同样遇到了如何在大尺度上开展实验研究的难题。他们通过文献分析,发现自20世纪80年代开始,全球有至少12个通过洲际或全国范围的多地点野外实验进行生态学研究的项目。他们认为这种大尺度和范围内的野外实验方法是有效解决上述难题的新操作规范。新操作规范被称作“协同分布式实验 ”(coordinated distributed experiments,CDEs),是一种“在全球的多个地点的多个研究团队并行开展的”野外实验操作规范,它具有“同时研究全球环境问题并且探索一般性生态学理论的优势,并提供受控实验内在的精确性”,认为这是“涌现出来的全球生态和环境科学假说验证的新工具”。他们认为自发出现的协同分布式实验的研究实践具有 5个属性(假说驱动的实验研究、多地理地点、标准化实验设计、标准化数据和协作的数据管理、知识产权共享)和3个特点(同步的数据收集、多研究团队、低预算)[28]。根据原文内容,推测属性是针对该规范的操作标准而言,特点则突出了该规范的优势。该文被《Frontiers in Ecology and The Environment》作为综述论文刊出,迅速受到了生态学界的关注。在Web of Science中,该文在1年半内被直接引用38次。其中,被引用最多的施引文献已被引用了30次,所有引用这篇施引文献的文献中被引用最多的达到了21次。以最多引用数的引文为线索进行文献分析,此文截止2015年8月25日已经发展出10层的前向引用,可见已经产生深远影响(图 1)。
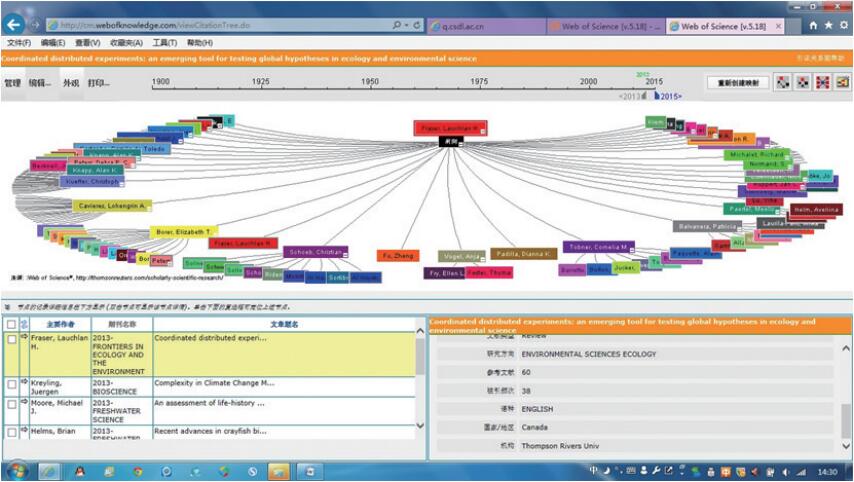
图1 Fraser等(2013)论文的前2层前向引用关系
Fig. 1 Top 2 tiers of front quotation relation of Fraser et al (2013)’s article
国内也有自发的协同分布式实验的应用。2011年启动的“中国大气气溶胶综合联网观测计划(CARE-China)”依托中国生态系统研究网络(CERN)在全国建立了36个研究型观测站点,统一仪器设备、观测标准和数据分析规范,协同展开了中国大气气溶胶及前体物联网观测研究,形成了中国区域大气气溶胶时间、空间、组分、浓度与光学特征变化的综合研究平台。该项研究由中国科学院大气物理研究所研究员王跃思主持,联合了中国科学院青藏高原研究所、地球环境研究所、广州地球化学研究所、北京化工大学和首都师范大学等7家研究单位。《美国气象学会会刊》(BAMS)主编对于研究成果的评价是:“首次进行了中国区域大气气溶胶物理、化学与光学特性综合监测,并尝试评估其在气候变化中的影响。 ”[29]
6 结论生态学假说必须经过严格的判决性实验验证才能成为理论。在经典的实验室实验和蓬勃发展的野外实验两种实验方法中,后者更适合作为生态学假说的判决性实验验证方法。但生态学野外实验的操作规范亟待完善,Fraser等2013年提出的CDEs操作规范有望解决这一难题,使得野外实验具有在全球的各个尺度和范围内验证生态学假说的逻辑可能和现实手段。
值得注意的是,CDEs仅仅是一个框架,Fraser等2013年的工作还远未完成,需要进一步完善并在此基础上进行实验以验证方法的有效性。
CDEs强调全球或大尺度和范围下开展多地点的、同时、同步、同规范的系列实验。理论上,如果在整个生态圈开展协同分布式实验是最佳的,但这不具备现实操作的可能性,也不合理。因此,实验地点的选取(数量与空间位置)成为被Fraser等2013年研究成果忽略的协同分布式实验的属性。虽然对于判决性实验是否存在是有争议的,但通过适当的限定、对具体问题的具体理论在某一时刻或某一时段进行野外实验,还是可成为有限结论的判决性实验。因此,目前不在全球尺度上(比如,群落及以下层次等级水平)提出的生态学假说可能更加适合进行协同分布式实验,针对全球尺度的生态学假说的协同分布式实验的属性、特点及具体操作标准的确定可能需要以前者长期的实践经验积累为前提。
即使协同分布式实验在全球尺度与范围内发展成熟,也仅仅是表明存在一般理论具备了逻辑可能,不代表一定能探寻到,也不能暗示具体的理论内容是什么,更不能确保未来不会出现反常现象而导致危机的出现并引发生态学革命。美国科学、工程和公共政策委员会在1995年曾指出:“科学成果必然是暂定的。科学家永远不能结论性地证明他们能完全精确地描述自然或物质世界的某些特性。 ”[8]
致谢 中国科学院大气物理研究所研究员王跃思、辛金元审阅了 CARE-China相关内容。
| [1] | 彭光华, 孙振钧, 吴文良 . 生态科学的内涵、本质与作用——纪念生态科学创立 140年[J]. 自然辩证法通讯, 2007(1): 53-59. |
| [2] | 斯通 . 如何在 Science上发表[R]. 北京: 中国科学院研究生院, 2010-11-1. |
| [3] | Brigandt I. Beyond reduction and pluralism: Toward an epistemology of explanatory integration in biology[J]. Erkenntnis, 2010, 73: 295-311. |
| [4] | Goldsmith. The way: An ecological world-view[M]. London: Rider, 1992. |
| [5] | Roughgarden Joan. Is there a general theory of community ecology[J].Biology and Philosophy, 2009, 24(4): 521-529. |
| [6] | Cooper G. Must there be a balance of nature[J]. Biology and Philosophy,2001, 16: 481-506. |
| [7] | Murray B G. Are ecological and evolutionary theories scientific[J].Biological Reviews. 2001, 76(2): 255-289. |
| [8] | 牛海山, 崔骁勇, 汪诗平, 等 . 生态学试验设计与解释中的常见问题[J]. 生态学报, 2009, 29(7): 3901-3910. |
| [9] | 林祥磊 . 生态学实验″伪重复″的方法论研究[D]. 北京: 中国科学院大学, 2013. |
| [10] | 林祥磊, 肖显静 .“生态学实验 ‘伪重复’”“真”“假”之辩[J]. 山西大学学报: 哲学社会科学版, 2013, 36(3):1-8. |
| [11] | 林祥磊 . 论生态学实验可重复性与伪重复的关联[J]. 科学技术哲学研究, 2016(1): 102-107. |
| [12] | Pacala S W, Rees M. Models suggesting field experiments to test two hypotheses explaining successional diversity[J]. The American Naturalist, 1998, 152(5): 729-737. |
| [13] | Hector A, Bazeley-White E, Loreau M, et al. Overyielding in grassland communities: Testing the sampling effect hypothesis with replicated biodiversity experiments[J]. Ecology Letters, 2002, 5: 502-511. |
| [14] | Srivastava D S, Kolasa J, Bengtsson J, et al. Are natural microcosms useful model systems for ecology[J]. Trends in Ecology and Evolution,2004, 19: 379-384. |
| [15] | Scheiner-Gurevitch. 生态学实验设计与分析[M]. 牟傅主, 译 . 北京:高等教育出版社, 2000. |
| [16] | 李博 . 生态学[M]. 北京: 高等教育出版社, 2000. |
| [17] | Svensson R, Lindegarth M, Siccha M, et al. Maximum species richness at intermediate frequencies of disturbance: consistency among levels of productivity[J]. Ecology, 2007, 88(4): 830-838. |
| [18] | Radford I, Cousens R. Invasiveness and comparative life-history traits of exotic and indigenous senecio species in australia[J]. Oecologia,2000, 125(4): 531-542. |
| [19] | Jaeschke A, Bittner T, Jentsch A, et al. The last decade in ecological climate change impact research: Where are we now[J].Naturwissenschaften, 2014, 101(1): 1-9. |
| [20] | 肖显静, 林祥磊 . 生态学实验的分类概况、欠缺及其完善[J]. 地理研究 . 2013(4): 653-662. |
| [21] | Holyoak M, Lawler P. The contribution of laboratory experiments on protists to understanding population and metapopulation dynamics[J].Advances in Ecological Research, 2005, 37: 245-271. |
| [22] | Griffen B, Drake J. A review of extinction in experimental populations[J]. Journal of Animal Ecology, 2008, 77(6): 1274-1287. |
| [23] | Quinn F, Dunham A E. On hypothesis testing in ecology and evolution[J]. American Naturalist, 1983, 122(5): 602-617. |
| [24] | Bender E, Case T, Gilpin M. Perturbation experiments in community ecology: Theory and practice[J]. Ecology, 1984, 65(1): 1-13. |
| [25] | Wu J, Loucks O. From balance of nature to hierarchical patch dynamics: A paradigm shift in ecology[J]. The Quarterly Review of Biology, 1995, 70(4): 439-466. |
| [26] | 沈泽昊 . 景观生态学的实验研究方法综述[J]. 生态学报, 2004, 24(4): 669-774. |
| [27] | Williams L, Auge H, Maron J. Testing hypotheses for exotic plant success: parallel experiments in the native and introduced ranges[J].Ecology, 2010, 91(5): 1355-1366. |
| [28] | Fraser L, Henry H, Carlyle C, et al. Coordinated distributed experiments:An emerging tool for testing global hypotheses in ecology and environmental science[J]. Frontier in Ecology and Environment Science, 2013, 11(3): 147-155. |
| [29] | 中国科学院大气物理研究所 .中国大气气溶胶联网观测研究获进展[EB/OL]. 2015- 08- 25[2015- 10- 03]. http://www.cas.cn/syky/201508/t20150819_4413172.shtml.2015-08-25. |