 2019, Vol. 39
2019, Vol. 39



2. 吉林建筑大学松辽流域水环境教育部重点实验室, 长春 130118;
3. 东北师范大学环境学院, 长春 130117
2. Key Laboratory of Songliao Aquatic Environment, Ministry of Education, Jilin Jianzhu University, Changchun 130118;
3. School of Environment, Northeast Normal University, Changchun 130117
冶金、印染、造纸等行业常产生大量的含Ca2+、Mg2+废水(周昌琴等, 2017), 这些硬度金属离子进入污水处理厂与活性污泥相接触, 影响活性污泥系统中的微生物群落系统组成、结构和功能.微生物群落结构与活性污泥的活性和絮凝沉降性紧密相关, 直接导致有机物去除率和脱氮效率的不同.因此, 研究硬度金属离子存在条件下微生物群落结构的演变规律对深入解析污染物的去除机制具有重要意义.
硬度金属离子(Ca2+、Mg2+)能够参与调节细胞内外渗透压、pH或氧化还原电位, 可以作为辅酶或激活酶的活性, 对微生物的生长代谢至关重要(杨红薇, 2014).许多学者研究了单一硬度金属离子对污泥性质的影响, 发现硬度金属离子能够促进活性污泥的生物絮凝性.Jiang等(2003)在SBR反应器中加入100 mg·L-1的Ca2+培养好氧颗粒污泥, 污泥颗粒更加致密, 沉降性提高, 多糖含量增加.Li等(2009)和Murthy等(1998)研究发现, 添加10 mg·L-1的Mg2+后, 缩短了SBR反应器污泥颗粒化的时间, 污泥颗粒更致密, 有利于污泥沉降、脱水和增加多糖含量.Yu等(2001)在UASB中引入浓度为150~300 mg·L-1的Ca2+后, 促进了生物量的累积, Ca2+通过吸附和粘附作用加速污泥的颗粒化过程.同时, 一些学者研究了单一硬度金属离子对微生物群落的影响, 例如, 周昌琴(2016)发现Ca2+的添加可改变反应器中优势菌所占比例, 但其对微生物多样性的影响较小.Schmidt等(1992)研究发现, 无Mg2+添加时, 微生物菌群主要由杆状产甲烷菌组成;添加100 mmol·L-1的Mg2+导致产甲烷菌群的解聚和大量单细胞的释放.Baodan等(2016)发现Tissieerella和Erysipelothrix分别在含有Na+和Ca2+的活性污泥系统中富集, 不同环境下微生物群落分布存在明显差异.也有学者研究了单一金属离子对系统稳定性和污染物去除能力的影响, 例如, 叶程晨(2016)发现, 当Ca2+为150 mg·L-1时, SBR工艺硝化菌和反硝化菌大量富集, 脱氮效率高.Liu等(2017)研究表明, 添加Mg2+(0.2 mol·L-1)导致污泥消化系统中与水解和酸化相关的细菌富集, 污泥脱水性能和除磷性能得到提高.Ye等(2016)在添加Ca2+(150 mg·L-1)的活性污泥中检测到黄杆菌和Rhodobacteraceae, 均能够分泌胞外聚合物, 促进微生物聚集, 并提高了污染物的去除率.这些研究主要探讨了单一金属离子的影响, 尚未系统研究共存金属离子的复杂体系对微生物群落结构和污染物去除效果的影响机制.
高通量测序技术可将成千上万个测序在一个平台同时进行, 在降低费用的同时, 还能够获得更加完整的DNA序列信息(Caporaso et al., 2010;王兴春等, 2012).本研究分别提取不同Ca2+/Mg2+物质的量比条件下活性污泥样品的总DNA, 并对其16S rDNA V4区域进行了PCR扩增和扩增产物的高通量测序分析, 结合有机污染物去除和脱氮效率的变化, 研究不同Ca2+/Mg2+物质的量比条件下微生物群落结构的差异性, 探索微生物脱氮机制.
2 材料与方法(Materials and methods) 2.1 试验装置接种的活性污泥取自吉林市污水处理厂.采用完全相同的3组SBR反应器(A、B和C组)进行实验, 反应器由有机玻璃制成, 有效容积为4 L, 反应器一侧设置3个取样口, 底部设置排泥口(图 1).好氧阶段采用空气泵曝气, 反应器底部设有曝气头, 通过转子流量计调节曝气强度, 控制DO为4~6 mg·L-1.SBR反应器每个运行周期由5个阶段组成:进水5 min→曝气4 h→搅拌2 h→沉降0.5 h→排水5 min→闲置1 h.利用机械搅拌保证缺氧阶段泥水充分混合.反应器运行温度控制在(20±2) ℃, 混合液悬浮固体浓度(mixed liquor suspended solids, MLSS)维持在2500 mg·L-1左右.每个运行周期进水3 L, 活性污泥沉降结束后排出相同体积的上清液.
 |
| 图 1 SBR反应器示意图 (1.电磁搅拌器;2.转子流量计;3.曝气泵;4.进水泵;5.曝气头;6.排泥阀;7.取样口;8.溶氧仪;9.pH计) Fig. 1 Schematic diagram of SBR |
实验用水采用模拟城市污水, 配水主要成分为:葡萄糖(0.28 g·L-1)、碳酸氢钠(0.172 g·L-1)、氯化铵(0.1 g·L-1)、磷酸二氢钾(0.02 g·L-1), 同时投加氯化钴(0.00042 g·L-1)、三氯化铁(0.00037 g·L-1)、钼酸钠(0.00015 g·L-1)、硫酸锰(0.00013 g·L-1)、硫酸铜(0.0001 g·L-1)等微量元素.通过投加CaCl2和MgSO4调控进水Ca2+/Mg2+物质的量比, 控制A、B和C组反应器进水Ca2+/Mg2+物质的量比分别为2、1和0.5(Ca2+浓度为1.1 mmol·L-1, Mg2+浓度分别为0.55、1.1和2.2 mmol·L-1).
2.2 水质分析项目和常规指标检测方法COD采用重铬酸钾法测定, NH4+-N采用纳氏试剂分光光度法测定, TN采用碱性过硫酸钾紫外分光光度法.混合液悬浮固体浓度(MLSS)采用重量法测定, SV30和污泥容积指数(sludge volume index, SVI)按标准方法测定.Zeta电位采用微电泳仪(上海中晨数字技术设备有限公司, JS94H2)检测, 污泥粒径采用激光粒度分析仪(LFC101, Ambivalue公司)测定.采用Wilén等(2003)提出的方法检测污泥絮凝能力(flocculation ability, FA).采用比耗氧呼吸速率(SOUR)表征活性污泥硝化活性, SOUR根据王建龙等(1999)提供的方法分析.
2.3 微生物群落结构分析 2.3.1 高通量测序在进水Ca2+/Mg2+比例分别为2、1和0.5的条件下, 分析SBR反应器的微生物群落结构.采用CTAB方法对样本的基因组DNA进行提取, 利用琼脂糖凝胶电泳检测DNA的纯度和浓度, 取适量的样本DNA于离心管中, 采用无菌水稀释至1 ng·μL-1.以稀释后的基因组DNA为模板, 对16S rDNA的V4区进行PCR扩增.采用New England Biolabs公司的Phusion® High-Fidelity PCR Master Mix with GC Buffer和高效高保真酶进行PCR, 确保扩增效率和准确性.反应条件:98 ℃预变性1 min, 然后30个循环(98 ℃变性10 s, 50 ℃退火30 s, 72 ℃延伸30 s), 最后72 ℃延伸5 min.采用Thermofisher公司的Ion Plus Fragment Library Kit 48 rxns试剂盒进行文库构建, 构建好的文库经过Qubit定量和文库检测合格后, 利用Thermofisher的Ion S5TMXL进行上机测序.
2.3.2 数据分析利用Uparse(v7.0.1001)软件进行聚类, 以97%的一致性将序列聚类为OTUs(Operational Taxonomic Units), 筛选OTUs中出现频数高的序列作为代表序列.采用MUSCLE(Version 3.8.31)软件进行快速多序列比对, 得到所有OTUs序列的系统发育关系, 进行Alpha多样性和Beta多样性分析.
采用Qiime软件(Version 1.9.1)计算Chao1、Shannon、Simpson、ACE指数, 利用R软件(Version 2.15.3)进行Alpha多样性指数组间差异性分析.
Chao1指数计算公式见式(1):
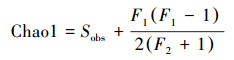
|
(1) |
式中, Sobs为测序分析得到的物种数, F1为样本中数量为1的数目, F2为样本中数量为2的数目.
ACE指数计算公式见式(2)~(3):
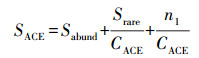
|
(2) |
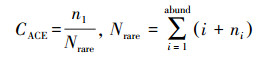
|
(3) |
式中, SACE为ACE指数, Srare为含有“abund”条序列或少于“abund”条序列的OTU数目, ni为含有i条序列的OTU数目, Sabund为多于“abund”条序列的OTU数目, abund默认为10.
Shannon指数计算公式见式(4):

|
(4) |
式中, H为Shannon指数, Pi为样品中属于第i种个体的比例.
Simpson指数计算公式见式(5):
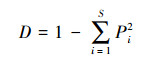
|
(5) |
式中, D为Simpson指数;S为物种数目.
3 结果与讨论(Results and discussion) 3.1 Ca2+、Mg2+共存对污染物去除率的影响Ca2+/Mg2+物质的量比对COD去除率的影响, 如图 2a 所示.在反应器运行0~15 d, 3组反应器的COD去除率均缓慢提高, 15 d后达到稳定.A、B和C组反应器COD去除率由88%分别增加至90%、91%和93%, 同时投加Ca2+、Mg2+对有机物的去除产生促进作用.经过30 d的运行, 3组反应器COD去除率大小顺序:C组>B组>A组.Mg2+体现出促进有机污染物降解的作用.
 |
| 图 2 Ca2+、Mg2+共存对污染物去除率的影响 Fig. 2 Effects of coexistence of Ca2+ and Mg2+ on the removal efficiencies of the pollutants |
Ca2+/Mg2+物质的量比对NH4+-N和TN的影响, 如图 2 b、2c所示.随着运行时间的增加, A、B和C组反应器NH4+-N去除率由74%增加至91%、93%和96%, TN去除率由44%分别增加至58%、62%和69%.3组反应器NH4+-N和TN去除率的大小顺序:C组>B组>A组.Mg2+比例增大促进了NH4+-N和TN去除率的提高.杨红薇等(2014)研究K+、Mg2+、Ca2+对污泥性能的影响时也发现, Mg2+对氨氮降解的促进能力高于Ca2+.本研究中, 随着进水Mg2+浓度的升高, TN去除率呈增大趋势, Mg2+促进了硝化菌和反硝化菌生长, 有效地提升了微生物的脱氮能力.
3.2 Ca2+、Mg2+共存对污泥性质的影响在不同Ca2+/Mg2+物质的量比下, 活性污泥SOUR的变化, 如图 3所示.随着进水Ca2+/Mg2+比例的降低, 污泥硝化活性增强.当进水Ca2+/Mg2+物质的量比为2时, SOUR、SOURAOB和SOURNOB分别为15.8、7.3和4.1 mg·g -1·h-1.当进水Ca2+/Mg2+为1时, 污泥比耗氧速率升高, SOUR、SOURAOB和SOURNOB分别为16.6、7.8和5.4 mg·g-1·h-1.当进水Ca2+/Mg2+物质的量比为0.5时, 污泥比耗氧速率继续增大, SOUR、SOURAOB和SOURNOB分别为18.4、10.2和6.9 mg·g -1·h-1, 氨氧化菌AOB和硝化菌NOB活性提高.同时还发现, 当进水Ca2+/Mg2+分别为2、1和0.5时, MLVSS/MLSS分别为0.57、0.61和0.71;随着Mg2+浓度增加, 促进了微生物的增长.Ca2+/Mg2+共存提高了SOURAOB和SOURNOB, 污泥硝化活性提高.
 |
| 图 3 不同Ca2+/Mg2+物质的量比条件下污泥比耗氧速率变化 Fig. 3 Variation of specific oxygen uptake rate of sludge under different molar ratios of Ca2+/Mg2+ |
在SBR反应器运行周期末, 分析3组反应器污泥混合液的粒径、FA和Zeta电位的变化.Ca2+/Mg2+对Zeta电位的影响, 如表 1所示.可以看出, 随着进水中Ca2+/Mg2+的下降, 污泥的Zeta电位呈上升趋势, 污泥絮体所吸附的正电荷增多.
| 表 1 不同Ca2+/Mg2+物质的量比条件下污泥混合液的Zeta电位 Table 1 Zeta potential of sludge mixed liquids at different Ca2+/Mg2+ molar ratio |
不同Ca2+/Mg2+物质的量比条件下污泥特性的变化, 如图 4所示.当进水Ca2+/Mg2+分别为2、1和0.5时, FA分别为27.4%、35.4%和46.0%.随着进水Ca2+/Mg2+的减小, 污泥重聚能力增加, 形成大而密实的污泥颗粒, 污泥粒径与FA呈增长趋势, 如图 4a所示.与单一投加Ca2+相比, Ca2+、Mg2+共存更有利于提高污泥的生物絮凝作用.图 4b表明, 随着进水Ca2+/Mg2+的降低, SV30与SVI呈减小趋势, 污泥沉降性增强.本研究中, 在进水Ca2+/Mg2+物质的量比为0.5的条件下, 污泥絮凝沉降性能最好.Mg2+能够促进蛋白质的合成, 并且在污泥絮状结构中与蛋白质结合;然而, Ca2+诱导碳水化合物的产生, 在胞外聚合物中选择性地与碳水化合物结合(Sanin et al., 2014).因此, Mg2+比例的增加有利于增强污泥絮体的结合能力, 提高污泥絮体的沉降性.
 |
| 图 4 不同Ca2+/Mg2+物质的量比条件下的污泥特性 Fig. 4 Sludge characteristics under different molar ratios of Ca2+/Mg2+ |
活性污泥系统中微生物多样性越高, 系统调节能力越强, 越有利于抵抗外界环境的变化, 保证工艺的稳定运行(Rowan et al., 2003;Hallin et al., 2005).Shannon指数和Simpson指数可以反映微生物群落的多样性, Shannon值越大, 微生物多样性越高.Chao1指数常用来估计物种总数, Chao1指数与ACE指数可以反映物种丰富度(张兰河等, 2018).不同Ca2+/Mg2+物质的量比对微生物群落多样性的影响, 如表 2所示.P2、P1和P0.5的Chao1指数和ACE指数远大于未添加硬度金属离子的污泥样品P0, Ca2+、Mg2+共存提高了微生物丰富度.同时, P2、P1和P0.5的Shannon指数和Simpson指数远大于P0, Ca2+、Mg2+共存增加了微生物多样性.随着进水Ca2+/Mg2+物质的量比的减小, Chao1指数和ACE指数增大, 微生物丰富度增大;Shannon指数下降, 微生物多样性降低;Simpson指数不变, 该区域常见物种未受到影响.
| 表 2 不同Ca2+/Mg2+物质的量比下微生物多样性 Table 2 Microbial diversity under molar ratios of Ca2+/Mg2+ |
采用主坐标分析(PCoA)和非度量多维尺度分析(NMDS)方法考察微生物群落的差异性, 两点间距离远近表示微生物差异性的大小(张兰河等, 2019).不同Ca2+/Mg2+物质的量比下微生物群落的差异性, 如图 5所示.进水Ca2+/Mg2+物质的量比的不同导致微生物群落之间具有显著差异.未添加硬度金属离子的微生物与Ca2+/Mg2+共存在时的微生物分别位于左右两侧, 这表明Ca2+、Mg2+共存的活性污泥系统微生物群落与未添加硬度金属离子活性污泥系统的微生物群落差异性显著.P1和P0.5的微生物群落之间的差异性小;与P1和P0.5相比, P2的微生物群落差异性大.
 |
| 图 5 微生物群落结构的差异性 (a.主坐标分析;b.非度量多维尺度分析;c.目水平聚类分析) Fig. 5 Differences of microbial community structure (a.Principal coordinate analysis; b.Non-metric multidimensional scaling analysis; c. Cluster analysis) |
根据OTU聚类和微生物丰度进行聚类分析, 结果如图 5c 所示.未添加硬度金属离子的微生物群落结构与硬度金属离子共存的微生物群落结构差异较大, P0的微生物群落产生分支, 这与Beta多样性分析结果相一致.图 2中污染物去除率的变化规律与微生物群落差异性的分析结果相符合, 这表明进水Ca2+/Mg2+物质的量比对微生物群落结构差异性产生影响, 从而导致系统的污染物去除率发生变化.
3.4 Ca2+、Mg2+共存对微生物群落结构的影响 3.4.1 门水平上微生物群落结构门水平上的微生物群落相对丰度, 如图 6a所示.在所有污泥样品中, 变形菌门均为优势菌门, 相对丰度最高.当进水Ca2+/Mg2+物质的量比分别为2、1和0.5时, 变形菌门、拟杆菌门、酸杆菌门相对丰度相近, 放线菌门相对丰度分别为14.4%、8.1%和7.5%, 厚壁菌门相对丰度分别为1.6%、3.4%和6.7%.与未添加金属离子的污泥样品相比, 当Ca2+、Mg2+共存时, 拟杆菌门和放线菌门的相对丰度增大.拟杆菌门不仅在水环境中广泛存在, 还是厌氧消化的主要菌门之一.拟杆菌门多为化能异养型细菌, 能够降解复杂的大分子有机物为小分子有机物(刘沛然, 2017).拟杆菌门丰度的增加, 增强了活性污泥体系对有机物的降解能力, 这与COD去除率的变化保持一致.
 |
| 图 6 微生物门(a)和目(b)水平物种相对丰度 Fig. 6 Relative abundance of microbial species at phylum (a) and order (b) level |
目水平上的微生物群落相对丰度的变化, 如图 6b所示.随着进水Ca2+/Mg2+物质的量比的降低, 黄单胞菌目(Xanthomonadales)一直是优势菌目, 其在分类学水平上属于变形菌门.与未添加硬度金属离子的污泥样品相比, Ca2+、Mg2+共存有利于Chitinophagales、Micrococcales和拟杆菌目的相对丰度增加.当进水Ca2+/Mg2+物质的量比分别为2、1和0.5时, Chitinophagales的相对丰度分别为7.3%、8.1%和8.3%, Micrococcales的相对丰度分别为7.0%、4.6%和4.3%, 拟杆菌目的相对丰度分别为5.5%、4.9%和4.4%.不同Ca2+/Mg2+物质的量化对微生物目水平上的群落丰度无明显影响.
3.4.3 属水平上微生物群落聚类分析图 7为属水平上的微生物聚类图, 大量参与脱氮过程的菌属被发现, 假单胞菌属Pseudomonas是典型的具有反硝化功能的微生物, Microbacterium是快速增长的专性好氧硝化菌属(Schmidt et al., 1992).当进水Ca2+/Mg2+物质的量比分别为2、1和0.5时, Niabella的相对丰度分别为1.53%、1.90%和1.95%, Dechloromonas的相对丰度分别为1.09%、0.79%和1.11%.Niabella是硝化细菌的代表性菌种(Wang et al., 2016), Dechloromonas是具有脱氮能力的代表性菌种(Yuan et al., 2016);与Ca2+/Mg2+物质的量比为2和1的进水条件相比, 当进水Ca2+/Mg2+物质的量比为0.5时, Niabella和Dechloromonas的相对丰度增大, 有利于提高反应器的硝化反硝化性能.
 |
| 图 7 微生物聚类图 Fig. 7 Microbial cluster graph |
1) 随着进水Ca2+/Mg2+物质的量比的降低, 微生物群落丰富度增大.进水Ca2+/Mg2+物质的量比为1和0.5的微生物群落之间的差异性较小;但它们与进水Ca2+/Mg2+物质的量比为2的微生物群落之间的差异性较大.
2) 随着进水Ca2+/Mg2+物质的量比的降低, 污泥硝化活性和絮凝沉降性增强;当进水Ca2+/Mg2+物质的量比为0.5时, 氨氮和总氮去除效果最好.同时投加Ca2+、Mg2+有助于提高SBR工艺的有机物降解能力和脱氮能力.
3) 变形菌门、拟杆菌门和放线菌门一直是Ca2+、Mg2+共存系统SBR工艺的优势菌门, 黄单胞菌目(Xanthomonadales)为优势菌目.大量参与脱氮过程的菌属在反应器内富集, 保证了良好的脱氮效果.
Caporaso J G, Kuczynski J, Stombaugh J, et al. 2010. QⅡME allows analysis of high-throughput community sequencing data[J]. Nature Methods, 7(5): 335–336.
DOI:10.1038/nmeth.f.303
|
Dilek Sanin F, Vatansever A, Turtin I, et al. 2006. Operational conditions of activated sludge:Influence on flocculation and dewaterability[J]. Drying Technology, 24(10): 1297–1306.
DOI:10.1080/07373930600838009
|
Hallin S, Lydmark P, Kokalj S, et al. 2005. Community survey of ammonia-oxidizing bacteria in full-scale activated sludge processes with different solids retention time[J]. Journal of Applied Microbiology, 99(3): 629–640.
DOI:10.1111/j.1365-2672.2005.02608.x
|
Jiang H L, Tay J H, Liu Y, et al. 2003. Ca2+ augmentation for enhancement of aerobically grown microbial granules[J]. Biotechnology Letters, 25(2): 95–99.
DOI:10.1023/A:1021967914544
|
Jin B D, Wang S Y, Xing L Q, et al. 2016. Long term effect of alkali types on waste activated sludge hydrolytic acidification and microbial community at low temperature[J]. Bioresource Technology, 200: 587–597.
DOI:10.1016/j.biortech.2015.10.036
|
Li X, Liu Q, Yang Q, et al. 2009. Enhanced aerobic sludge granulation in sequencing batch reactor by Mg2+ augmentation[J]. Bioresource Technology, 100(1): 64–67.
DOI:10.1016/j.biortech.2008.06.015
|
Liu C C, Huang C, Sun X Y, et al. 2017. The effect of Mg2+ on digestion performance and microbial community structures in sludge digestion systems[J]. Environmental Science and Pollution Research, 24(21): 17474–17484.
DOI:10.1007/s11356-017-9320-y
|
刘沛然. 2017. Ca2+、Mg2+对垃圾渗滤液厌氧生物处理及微生物群落的影响研究[D].合肥: 安徽建筑大学
|
Murthy S N, Novak J T, Haas R D D. 1998. Monitoring cations to predict and improve activated sludge settling and dewatering properties of industrial wastewaters[J]. Water Science and Technology, 38(3): 119–126.
DOI:10.2166/wst.1998.0189
|
Rowan A K, Snape J R, Fearnside D, et al. 2003. Composition and diversity of ammonia-oxidising bacterial communities in wastewater treatment reactors of different design treating identical wastewater[J]. FEMS Microbiology Ecology, 43(2): 195–206.
DOI:10.1111/j.1574-6941.2003.tb01059.x
|
Schmidt J E, Macario A J L, Ahring B K, et al. 1992. Effect of magnesium on methanogenic subpopulations in a thermophilic acetate-degrading granular consortium[J]. Applied and Environmental Microbiology, 58(3): 862–868.
|
王建龙, 吴立波, 齐星, 等. 1999. 用氧吸收速率(OUR)表征活性污泥硝化活性的研究[J]. 环境科学学报, 1999, 19(3): 225–229.
DOI:10.3321/j.issn:0253-2468.1999.03.001 |
王兴春, 杨致荣, 王敏, 等. 2012. 高通量测序技术及其应用[J]. 中国生物工程杂志, 2012, 32(1): 109–114.
|
Wang X Y, Xing D F, Ren N Q. 2016. p-Nitrophenol degradation and microbial community structure in biocathode bioelectrochemical system[J]. RSC Advances, 6(92): 89821–89826.
DOI:10.1039/C6RA17446A
|
Wilén B M, Jin B, Lant P. 2003. The influence of key chemical constituents in activated sludge on surface and flocculating properties[J]. Water Research, 37(9): 2127–2139.
DOI:10.1016/S0043-1354(02)00629-2
|
杨红薇, 陈佼, 张建强. 2014. K+、Ca2+、Mg2+对高盐肝素废水处理的影响[J]. 环境工程学报, 2014, 8(10): 4267–4272.
|
Ye C, Yang X, Zhao F J, et al. 2016. The shift of the microbial community in activated sludge with calcium treatment and its implication to sludge settleability[J]. Bioresource Technology, 207(2016): 11–18.
|
叶程晨. 2016. Ca(Ⅱ)介导的活性污泥微生物种群结构变化对活性污泥沉降性能的影响[D].南京: 南京农业大学
http://cdmd.cnki.com.cn/Article/CDMD-10307-1017259740.htm |
Yu H Q, Tay J H, Fang H H P. 2001. The Roles of Calcium in Sludge Granulation During UASB Reactor Start-up[J]. Water Research, 35(4): 1052–1060.
DOI:10.1016/S0043-1354(00)00345-6
|
Yuan Y, Liu J J, Ma B, et al. 2016. Improving municipal wastewater nitrogen and phosphorous removal by feeding sludge fermentation products to sequencing batch reactor (SBR)[J]. Bioresource Technology, 222: 326–334.
DOI:10.1016/j.biortech.2016.09.103
|
张兰河, 田蕊, 陈子成, 等. 2018. NaCl盐度对A2/O工艺去除废水污染物和系统微生物的影响[J]. 农业工程学报, 2018, 34(10): 231–237.
DOI:10.11975/j.issn.1002-6819.2018.10.029 |
张兰河, 郑晶, 田蕊, 等. 2019. Na+和K+共存对A2/O工艺脱氮除磷效果及污泥性质的影响[J]. 农业工程学报, 2019, 35(11): 206–213.
DOI:10.11975/j.issn.1002-6819.2019.11.024 |
周昌琴. 2016.高钙废水生物处理过程中微生物群落及其生物酶的变化[D].兰州: 兰州理工大学
http://cdmd.cnki.com.cn/Article/CDMD-10731-1016906492.htm |
周昌琴, 孔秀琴, 陈磊, 等. 2017. PESA对高钙废水中污泥酶活性的影响[J]. 环境工程学报, 2017(4): 2212–2218.
|