 2018, Vol. 38
2018, Vol. 38



抗生素被长期用于预防和治疗人类和动物疾病, 并在动物养殖业当中广泛使用, 主要作为生长促进剂以亚治疗剂量加入动物饲料中(周启星等, 2007).大多数抗生素只有15%可以被吸收利用, 剩余的通过尿液和粪便排入自然生态系统(Zhang et al., 2011).有研究表明, 在2013年, 中国大约有53800 t抗生素释放到河流和水道中(Zhang et al., 2015).进入到环境中的抗生素除了直接对环境造成污染外, 还可能诱导环境中耐药性细菌和抗生素抗性基因(Antibiotic Resistance Genes, ARGs)的产生(Kim et al., 2004).抗生素的广泛生产和滥用促进了ARGs在环境中的富集和传播, 2010年发现的超级细菌, 其携带耐药基因NDM-1编码的β-内酰胺酶有消解大多数抗生素的能力(Zhang et al., 2011).
国内外学者已对地下水、地表水、污水处理厂废水、饮用水中抗生素和ARGs的含量展开研究.在我国的珠江、黄浦江、黄河、海河流域中都检测到抗生素与ARGs, 例如, Lin等(2015)对长江入海口及沿海沉积物中的ARGs展开研究, 发现sul类、tet类、intⅠ基因在这一区域内普遍存在, 含量从河口到海岸呈现出下降的趋势;Jiang等(2013)以黄浦江流域和流域内饮用水水源地为研究目标, 发现水源地ARGs的含量高于黄浦江流域的其他地区;Huerta等(2013)对水源地水库抗生素、抗性基因与微生物群落之间的关系进行了研究.
鉴于国内外现阶段研究目标聚焦在单一水环境, 针对ARGs从河流系统到水库系统赋存特征的研究较少, 且夏季时期藻类含量高, 微生物活跃, 是水库水质的高风险时期.因此, 本研究选择夏季时期长江下游某水源型水库为研究对象, 分别采集水样和沉积物样本并使用高通量定量PCR和实时荧光定量PCR方法, 检测水体和沉积物中的八大类共285种目前已知的主要类型抗生素抗性基因, 并对其赋存分布特征进行分析, 以期为水库ARGs污染控制及水质安全保障提供基础依据.
2 材料与方法(Materials and methods) 2.1 样品采集该水源型水库位于长江下游(31°48'N, 121°57'E), 供水规模超过719万m3·d-1.本研究于2016年5月和6月赴水库采样, 围绕整个水库共设置4个采样点(图 1).
 |
| 图 1 水库地理位置示意图 Fig. 1 Location of reservoir |
S1样点位于长江下游(水库外), 是水库的入水口, 入水口水流速度和浊度变化明显(5、6月样品分别命名为5-S1、6-S1);S2样点位于水库内下游水闸, 该泵站具有维持水库蓄水量恒定的功能(5、6月样品分别命名为5-S2、6-S2);S3样点位于库内输水泵站, 该泵站将水库内水输送至自来水厂(5、6月样品分别命名为5-S3、6-S3);S4样点位于库内中游位置, 是水库重要的一个巡视位置(5、6月样品分别命名为5-S4、6-S4).在S1~S4样点距离水面0.5 m处采集10 L水样, 立即运回实验室采用0.22 μm滤膜进行过滤并于-80 ℃冰箱保存;在库内中游处用抓泥斗采集表层沉积物, 命名为SE4(5、6月样品分别命名为5-SE4、6-SE4), 采取的沉积物立即用铝箔纸封存并于-80 ℃冰箱保存.
2.2 水样及沉积物DNA提取水样和沉积物的DNA分别用E.Z.N.A.® Water DNA Kit和E.Z.N.A.® Soil DNA Kit (OMEGA, USA)提取, 根据试剂盒的说明书, 按操作步骤进行提取.提取出的DNA用1%琼脂糖凝胶电泳和Thermo Nanodrop2000微量分光光度计(Thermo Scientific, USA)进行检测, DNA样品用QuantiFluor® dsDNA System (Promega Corporation, USA)试剂盒测定双链DNA浓度, 根据测定的样品DNA浓度, 用灭菌的超纯水统一稀释至30 ng·μL-1.
2.3 高通量荧光定量PCR使用SmartChip Real-Time PCR Systems(WaferGen Inc., USA)高通量荧光定量反应平台测定DNA样品的ARGs.该SmartChip纳米孔平台可用于大规模基因表达研究, 每次运行可处理5184个纳米孔反应(Zhu et al., 2013).使用的293对引物在其他研究者的研究中已经被有效验证, 新加入3对引物, 1对用于测定超级细菌抗生素抗性基因blaNDM-1, 1对用于测定intⅠ1整合子基因(class1 intergon), 1对用于测定CintⅠ1临床医学意义上的整合子基因(clinical class1 intergon)(黄福义等, 2016).
每个纳米孔用于PCR扩增反应的总体积为100 nL, 由1×LightCycler 480 SYBR® GreenⅠMaster Mix(Roche, USA)、1 mg·mL-1牛血清蛋白(BSA)、nuclease-free PCR-grade water、500 nmol·L-1上下游引物和5 ng·μL-1 DNA模板组成.使用SmartChip Multisample Nanodispenser (MSND)将PCR混合物释放到芯片的纳米孔中, 随后用SmartChip Cycler进行实时qPCR.每个芯片都有无模板的阴性对照.反应条件:初始95 ℃下预变性10 min;95 ℃变性30 s, 60 ℃退火延伸30 s, 40个循环.仪器程序自动升温进行熔解曲线分析, 导出qPCR反应得到的扩增效率介于1.8~2.2的数据.根据该系统的精确度和灵敏度, 将仪器的检测限设为循环次数CT=31(Wang et al., 2014).
2.4 实时荧光定量PCR使用PikoReal-Time PCR System(Thermo Scientific., USA)进行qPCR定量分析.通过TA克隆, 构建带有pMD® 18-T Vetor (TaKaRa Biotechnology, China)的质粒载体, 将10倍浓度梯度稀释的质粒作为定量的标准品, 用于制作绝对定量的标准曲线.qPCR反应体系如下:12.5 μL SYBR® Premix Ex TaqTM Ⅱ(2×) (TaKaRa Biotechnology, China), 正向、反向引物(10 μmol·L-1)各0.5 μL, DNA模板(10 ng·μL-1)1 μL, 总反应体积为25 μL.qPCR反应条件为:95 ℃下预变性30 s;95 ℃变性5 s, 60 ℃退火30 s, 循环40次;72 ℃延伸3 min.
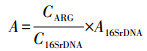
2.5 数据分析根据Looft等(2012)的研究, 用公式(1)、(2)分别计算基因的相对拷贝数和绝对丰度.
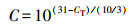
|
(1) |
|
(2) |
式中, C是基因相对拷贝数;CT是PCR反应收集到特定荧光时的循环次数;A是基因的绝对丰度(copies·L-1或copies·g-1).
利用Microsoft Excel 2010进行计算分析, 利用SPSS 21.0进行相关性分析, 利用Origin Pro 9.0做图, Circos图利用Circos software (http://circos.ca/)制作, 热图利用MeV制作.
3 结果与讨论(Results and discussion) 3.1 水库抗生素抗性基因多样性按照抗性基因对应的抗生素类型将295种ARGs分为九大类抗生素抗性基因(黄福义等, 2016), 分别为磺胺类抗生素(Sulfonamide)抗性基因(7种)、四环素类抗生素(Tetracycline)抗性基因(39种)、氯霉素类抗生素(Chloramphenicol)抗性基因(3种)、β-内酰胺类抗生素(β-Lactamase)抗性基因(54种)、大环内脂类-林肯酰胺类-链阳性菌素B类抗生素(MLSB)抗性基因(45种)、氨基糖苷类抗生素(Aminoglycoside)抗性基因(36种)、万古霉素类抗生素(Vancomycin)抗性基因(32种)、多重抗药类抗生素(Multidrug)抗性基因(52种)和其他类抗生素(Others)抗性基因(27种).以上九大类ARGs均在水体和沉积物中检测到.
如图 2所示, 5-S1样品检出104种ARGs, 5-S2样品检出118种, 5-S3样品检出115种, 5-S4样品检出118种, 5-SE4样品检出122种;6-S1样品检出101种ARGs, 6-S2样品检出116种, 6-S3样品检出114种, 6-S4样品检出117种, 6-SE4样品检出124种.有研究人员对水体和沉积物中的抗性基因污染情况展开了研究, 例如, Xu等(2016)检测了钱塘江流域下游饮用水厂进水、出水和自来水中ARGs的种类, 分别检出168、115、117种ARGs;Wang等(2014)研究了公园沉积物中ARGs的分布情况, 选取北京、石家庄、济南、武汉、昆明、包头和乌鲁木齐地区7个使用再生水灌溉植物的公园, 结果显示, 7个公园共检出147种ARGs.Zhu等(2017)对辽宁、河北、天津、浙江、福建、广东、广西共18个河口沉积物的ARGs开展研究, 各河口检测到的ARGs种类为85~158种, 平均为118.与这些研究结果相比, 长江下游及其蓄水水库的抗性基因污染程度处于中等水平.
 |
| 图 2 研究区不同样点检测到的抗生素抗性基因种类数 Fig. 2 Number of detected antibiotic resistance genes in different sampling sites |
ARGs的相对丰度表明, β-内酰胺类、多重抗药类、其他类(包含转座子基因、整合子基因)抗生素抗性基因在样品中的相对丰度较高(图 3).β-内酰胺类ARGs在10个样品中的相对丰度分别为4.58×10-3(5-S1)、7.53×10-2(5-S2)、6.26×10-2(5-S3)、7.74×10-2(5-S4)、5.68×10-2(5-SE4)、3.25×10-2(6-S1)、6.85×10-2(6-S2)、3.55×10-2(6-S3)、5.70×10-2(6-S4)、5.12×10-2(6-SE4).本实验主要研究bla和ampC两类基因, 实际检测到5种ampC基因, 相对丰度为7.48×10-6~7.43×10-4, ampC-06只在沉积物中检出;14种bla基因的相对丰度为1.05×10-6~1.08×10-2, 其中, blaPAO、blaSHV-02只在沉积物中检出, blaVEB只在水体中检出.在澳大利亚的饮用水及医院和城市废水中都检测到ampC, 德国的废水中该ARG检出率为78% (Schwartz et al., 2003).bla通常在动物来源的病原体(气单胞菌、肠杆菌、沙门氏菌)中被检测到, 在地表水中同样被发现(Zhang et al., 2009).β-内酰胺类抗生素具有低毒性的特点, 在人类医疗和农耕养殖业中被广泛使用, 人类使用的抗生素药物(青霉素、头孢菌素)中有2/3都为这一大类, 也是德国兽药用量最大的一类抗生素(Stoll et al., 2012).因此, 该类ARGs的高相对丰度与检出率可能是由于医疗废水、生活污水、养殖废水排放进入环境所导致.
 |
| 图 3 水体和沉积物中ARGs相对丰度(a.5月,b.6月) Fig. 3 Relative abundance of ARGs in water and sediment samples |
多重抗药类ARGs的相对丰度较高(4.66×10-3~9.55×10-2), 5-S1处相对丰度为4.66×10-3, 5-S4处为4.04×10-2, 5-SE4处为7.56×10-2, 在水库中的最大富集倍数(基因相对拷贝数的比值)为9.5, 沉积物中为1.87;6月库外相对丰度为3.53×10-2, 最高相对丰度出现在S2处, 为9.55×10-2, 6-SE4处为9.30×10-2, 在水库中的最大富集倍数为2.7, 沉积物中为2.0.这类抗生素在医院和养殖业中被广泛使用, 残余的抗生素会诱导抗性病菌和多重抗药基因产生, 九龙江流域中有70.6%的细菌菌株对抗生素呈现多重耐药性.有研究发现, 废水经过污水处理厂, 多重耐药性水平提高(Wang et al., 2014), 这也可能是多重抗药类ARGs进入水体的一个重要途径.中国抗生素耐药性增长速度很快, 因此, 合理使用抗生素, 避免多重耐药病原体在环境中的传播显得尤为重要.
氯霉素类ARGs相对丰度与其他几类ARGs相对丰度相比处于中等污染水平(8.24×10-3~2.10×10-2).这可能是因为从20世纪50年代起, 该类抗生素就被长时间应用于治疗动物和人类疾病.尽管现如今很多国家都禁止在家禽中使用该类抗生素, 但因为它具有抗生素效力稳定、价格低廉的特点, 在中国水产养殖和畜牧业中仍部分存在非法使用该类抗生素的现象, 因此, 这类抗生素对应的ARGs在环境中仍有存在.
其他类抗生素抗性基因包括转座子基因、整合子基因和喹诺酮类基因, 转座子和整合子可以捕获整合及表达抗性基因.在下游水闸采样点处(S2)相对丰度高达9.24×10-2, 高于S4处相对丰度(2.11×10-2).本研究发现, 8个水体样品测得的CintⅠ-1(class 1)、intⅠ-1(clinical)的相对含量与ARGs总量之间存在显著相关性(分别为r=0.750, p<0.05;r=0.971, p<0.05).在城市表层水中分离的弧菌菌株中发现, CintⅠ-1(class1)和甲氧苄氨嘧啶、氨基糖苷、β-内酰胺形成具有抗性的共轭原件(Zhang et al., 2009).Zhu等(2017)研究发现, ARGs的丰度与整合子、转座子之间分别存在显著相关性(p<0.01).Luo等(2011)研究发现, 海河沉积物中intⅠ和sulⅠ相对含量之间有显著相关性, 并且证明intⅠ参与了环境中sulⅠ的传播.一些研究表明, intⅠ-1可以通过基因盒整合, 表达超过100种ARGs(包括氨基糖苷类类、β-内酰胺类、磺胺类)(Gillings et al., 2008), 这说明CintⅠ-1(class 1)、intⅠ-1(clinical)可能对ARGs的水平传播起到关键作用.
3.3 抗生素抗性基因的分布特征图 4显示了检测到的ARG相对丰度在每个采样点的分布特征.比较发现, 库外ARGs种类与相对丰度均小于库内, 其中, β-内酰胺类ARGs在库中富集了4.75~16.89倍, 多重抗药类ARGs在库中富集了3.61~9.51倍, 其他类ARGs在库中富集了2.18~5.05倍;比较S4与SE4发现, 沉积物的ARGs种类多于水体(SE4>S4), β-内酰胺类ARGs在沉积物中富集了0.73~1.58倍, 沉积物中氯霉素类、多药类和万古霉素类ARGs相对丰度高于水体, 说明ARGs在沉积物中得到富集.mexF、intⅠ-1(clinic)、cphA-01是3种优势抗性基因, 其中, intⅠ-1(clinic)是对ARGs水平迁移有关键作用的整合子基因.5-S1与6-S1聚类, 5-S3与6-S3聚类, 5-S4与6-S4聚类, 5-SE4与6-SE4聚类, 库外采样点(S1)与库内3个点(S2、S3、S4)聚类, 沉积物与水样聚类, 表明水库沉积物的抗性分布格局不同于水库内和水库内的3个水样.
 |
| 图 4 水体和沉积物中ARGs分布格局 Fig. 4 Profile of ARGs in water and sediment samples |
有12种ARGs (erm(34)、acrA-02、acrA-03、acrF、adeA、mexA、yceL_mdtH-01、vanC2_vanC3、vanHB、tnpA-05、aac、blaVIM)只在水库内检出, 库内ARGs多样性大于库外.而本课题组前期研究发现, 水库对总氮、总磷等常规指标, 以及药品和个人护理用品类污染物(PPCPs), 如咖啡因、双酚A、抗生素等有削减作用, 并未出现库内ARGs丰度高于库外的现象.出现这种差异的原因可能是ARGs作为一种生物特性的新型污染物, 有别于传统的化学特性的新型污染物, 其生物体间具有的传播机制和独特的环境行为尚需进一步的研究.但已有研究表明, ARGs丰度会随着采样点处流速的增大而减小(Jiang et al., 2013).而本研究中水库上游进水闸(S1)流速(0.2 m·s-1)>库区流速(0.01~0.19 m·s-1)>下游水闸(S2)流速(0.01~0.03 m·s-1)(袁建忠等, 2012), 显示长江下游的水流速度明显高于库内流速.此外, 16S rDNA绝对定量结果显示, 从库外S1到库内S2、S3、S4及SE4的16S rDNA的均值分别为4.79×108、1.13×1010、1.89×1010、4.18×1010、6.56×109 copies·g-1.表明库内及其沉积物中的细菌数量显著高于库外1~2个数量级, 而丰富的生物量有助于ARGs在不同细菌间的转移和积累.因此, 水流速度和细菌数量可能是造成这种现象的主要原因.
10种ARGs只在沉积物中被检出, 分别为vgaA-01、tetM-01、tetPB-01、tetS、bacA-02、catB3、aacC1、ampC-06、blaPAO、blaSHV-02.环境中残留的抗生素可能会诱导ARGs的出现, 且抗生素特性的差异会影响ARGs在环境中的分布.大环内酯类抗生素易被水解且吸附于沉积物中, 喹诺酮类抗生素同样易被光降解和吸附于沉积物中(Luo et al., 2011), 这两种抗生素对应的ARGs在相对丰度与种类上均显示出沉积物大于水体的特征.这说明这类易水解、光解且吸附于沉积物的抗生素对应的ARGs在水体中的相对丰度与种类均小于对应沉积物.以上结果显示, 沉积物介质具有选择性富集大环内酯类和喹诺酮类抗性基因的功能, 也进一步证实了水体介质与沉积物介质之间ARGs分布特征存在差异.
3.4 抗生素抗性基因的绝对丰度每一大类ARGs和每个样品中ARGs绝对丰度和分布特征见图 5.图 5显示, 从6-S1到5-SE4样品, ARGs的总含量依次为1.60×108、1.31×108、1.12×109、2.70×109、2.53×109、3.50×109、5.40×109、6.30×109 copies·L-1和1.80×109、7.86×108 copies·g-1.
 |
| 图 5 每种ARG在5个采样点中的分布 Fig. 5 Distribution of each ARG type in five Sampling points |
氨基糖苷类(36种)抗性基因在水样中的绝对丰度总和为4.22×108 copies·L-1, 其中, 6-S1为4.85×106 copies·L-1, 5-S1为6.16×106 copies·L-1, 6-S2为3.62×107 copies·L-1, 5-S2为3.57×107 copies·L-1, 6-S3为5.98×107 copies·L-1, 5-S3为5.99×107 copies·L-1, 6-S4为1.17×108 copies·L-1, 5-S4为1.03×108 copies·L-1;在沉积物中的绝对丰度总和为4.45×107 copies·g-1, 其中, 6-SE4为3.44×107 copies·g-1, 5-SE4为1.02×107 copies·g-1.其他各类ARGs绝对丰度分布度情况如图 5所示.
β-内酰胺类(54种)抗性基因检出率为44%, 水体中浓度为3.44×107~2.49×109 copies·L-1, 沉积物中含量分别为2.36×108 copies·g-1和4.59×108 copies·g-1.其中, blaTEM绝对丰度为6.68×104~1.96×107 copies·L-1, blaCARB、blaSHV、blaSHV-5未被检出.Jiang等(2013)在黄浦江流域检测到blaTEM浓度为1.4×106~2.0×107 copies·L-1, 是最高浓度S4处(1.46×107 copies·L-1)的0.10~1.37倍;Hu等(2008)研究发现, 超过95%的环境样本中检测到blaTEM, 而blaCARB、blaSHV、blaSHV-5的检出率低于1%.该类ARGs在本水库的检出情况与大部分环境样本一致, 且blaTEM含量低于黄浦江流域.
研究结果显示, 磺胺类ARGs和四环素类ARGs绝对丰度不高, 但这两类ARGs在环境中广泛传播(Gillings et al., 2008).sul Ⅰ、sul Ⅱ在本实验中的检出率均为100%, 磺胺类ARGs的含量高于四环素类ARGs的含量, 这一结果与黄浦江流域研究结果一致(沈群辉等, 2012).dfrA1浓度为0~7.95×106 copies·L-1, dfrA12浓度为0~2.83×105 copies·L-1, dfrA12检出率较低, 分布特征与在德国、澳大利亚的研究结果一致(Stoll et al., 2012).这两种ARGs在环境中的分布特征可能是由于sul Ⅰ、sul Ⅱ属于在环境中比较常见的二氢叶酸合成酶基因, 而dfrA1、dfrA12属于在环境中比较少见的二氢叶酸还原酶基因, 这使得sul Ⅰ、sul Ⅱ基因被广泛检出.
11种四环素类(39种)抗性基因被检出, 其中, tetA-02、tetD-02、tetG-01、tetG-02、tetL-02、tetPB-02、tetR-02检出率为100%, 水体中tetA-02浓度为5.98×104~1.20×107 copies·L-1, 低于在太湖中的浓度(107~108 copies·L-1), tetM-01、tetPB-01、tetS只在沉积物中检出.Zhang等(2011)在中国和其他地区发现, 13种四环素类ARGs中, tetG的丰度最高, 其次是tetA、tetC.Hu等(2008)在自然水体中检测了20种四环素类ARGs, 结果只检测到tetA、tetB、tetM 3种.这可能是由于tetA、tetB、tetG由可迁移的载体, 如转座子和较小的质粒携带, 而质粒具有移动性较强的特点, 因此, 可以在耐药细菌之间稳定传播, 而tetO、tetS普遍存在于不可移动的大的共轭质粒或染色体中, 所以这类基因转移和传播的可能性较低(Schwartz et al., 2003).从S1到S4 ARGs的绝对丰度依次递增, 绝对丰度最高的3类ARGs为多重抗药类、β-内酰胺类、其他类ARGs.本水库与其他地区磺胺类和四环素类检出率都较高, 但长江下游和水库内ARGs浓度与其他流域相比较低.
4 结论(Conclusions)1) 夏季时期长江下游某水源型水库平均检测到118种ARGs, 与我国其他地区相比, 检出ARGs种类数处于中等水平.
2) 九大类ARGs中, β-内酰胺类、多药类、氯霉素类和其他类ARGs相对丰度与绝对丰度均高于其他五大类;四环素类ARGs和磺胺类ARGs绝对丰度低于其他几大类ARGs, 但因为sulⅠ、sulⅡ在环境中常见, tetA、tetG由移动性强的质粒携带, 检出率高于其他ARGs.
3) 移动性强的ARGs在环境中检出频率较高, 水体中整合子CintⅠ-1(class 1)、intⅠ-1(clinical)与ARGs总量之间存在显著的正相关性, 具有促进ARGs水平转移的作用, 是ARGs传播的重要途径.
4) 夏季时期ARGs的分布特征受环境介质影响, 使得水库内外, 以及水体与沉积物之间ARGs分布存在差异.低流速环境易于微生物生长, 细菌多样性多, 使得ARGs易在不同细菌间传播;水库内检测到的ARGs种类和丰度均大于长江下游, 这说明缓流水体易成为ARGs高风险区.
Gillings M, Boucher Y, Labbate M, et al. 2008. The evolution of class 1 integrons and the rise of antibiotic resistance[J]. Journal of Bacteriology, 190(14): 5095–5100.
DOI:10.1128/JB.00152-08
|
Huerta B, Marti E, Gros M, et al. 2013. Exploring the links between antibiotic occurrence, antibiotic resistance, and bacterial communities in water supply reservoirs[J]. Science of the Total Environment, 456: 161–170.
|
黄福义, 李虎, 安新丽, 等. 2016. 城市生活污水和生活垃圾渗滤液抗生素抗性基因污染的比较研究[J]. 环境科学, 2016, 37(10): 3949–3954.
|
Hu J, Shi J, Chang H, et al. 2008. Phenotyping and genotyping of antibiotic-resistant Escherichia coli isolated from a natural river basin[J]. Environmental Science & Technology, 42(9): 3415–3420.
|
Jiang L, Hu X L, Xu T, et al. 2013. Prevalence of antibiotic resistance genes and their relationship with antibiotics in the Huangpu River and the drinking water sources, Shanghai, China[J]. Science of the Total Environment, 4(38): 267–272.
|
Kim S R, Nonaka L, Suzuki S. 2004. Occurrence of tetracycline resistance genes tet (M) and tet (S) in bacteria from marine aquaculture sites[J]. FEMS Microbiology Letters, 237(1): 147–156.
DOI:10.1111/fml.2004.237.issue-1
|
Lin L, Liang X M, Chen X, et al. 2015. Occurrences and distribution of sulfonamide and tetracycline resistance genes in the Yangtze River Estuary and nearby coastal area[J]. Marine Pollution Bulletin, 8(36): 304–310.
|
Luo Y, Xu L, Rysz M, et al. 2011. Occurrence and transport of tetracycline, sulfonamide, quinolone, and macrolide antibiotics in the Haihe River Basin, China[J]. Environmental Science & Technology, 45(5): 1827–1833.
|
Looft T, Johnson T A, Allen H K, et al. 2012. In-feed antibiotic effects on the swine intestinal microbiome[J]. Proceedings of the National Academy of Sciences of the United States of America, 109(5): 1691–1696.
DOI:10.1073/pnas.1120238109
|
苏建强, 黄福义, 朱永官. 2013. 环境抗生素抗性基因研究进展[J]. 生物多样性, 2013, 21(4): 481–487.
|
Schwartz T, Kohnen W, Jansen B, et al. 2003. Detection of antibiotic-resistant bacteria and their resistance genes in wastewater, surface water, and drinking water biofilms[J]. FEMS Microbiol Ecology, 43(3): 325–335.
DOI:10.1111/fem.2003.43.issue-3
|
Stoll C, Sidhu J P S, Tiehm A, et al. 2012. Prevalence of clinically relevant antibiotic resistance genes in surface water samples collected from Germany and Australia[J]. Environmental Science & Technology, 46(17): 9716–9726.
|
沈群辉, 冀秀玲, 傅淑珺, 等. 2012. 黄浦江水域抗生素及抗性基因污染初步研究[J]. 环境科学, 2012, 21(10): 1717–1723.
|
Wang F H, Miao M, Su J Q, et al. 2014. High throughput profiling of antibiotic resistance genes in urban park soils with reclaimed water irrigation[J]. Environmental Science & Technology, 48(16): 9079–9085.
|
Xu L, Ouyang W Y, Qian Y Y, et al. 2016. High-throughput profiling of antibiotic resistance genes in drinking water treatment plants and distribution systems[J]. Environmental Pollution, 213: 119–126.
DOI:10.1016/j.envpol.2016.02.013
|
袁建忠, 李杰. 2012. 青草沙水库库区流态及淤积分布特征研究[J]. 城市道桥与防洪, 2012(9): 181–183.
|
周启星, 罗义, 王美娥. 2007. 抗生素的环境残留、生态毒性及抗性基因污染[J]. 生态毒理学报, 2007, 2(3): 243–251.
|
Zhang D D, Lin L F, Luo Z X, et al. 2011. .Occurrence of selected antibiotics in Jiulongjiang River in various seasons, South China[J]. Journal of Environmental Monitoring, 13(7): 1953–1960.
DOI:10.1039/c0em00765j
|
Zhang Q Q, Ying G G, Pan C G, et al. 2015. Comprehensive evaluation of antibiotics emission and fate in the river basins of China:source analysis, multimedia modeling, and linkage to bacterial resistance[J]. Environ Environmental Science & Technology, 49(11): 6772–6782.
|
Zhu Y G, Johnson T A, Su J Q, et al. 2013. Diverse and abundant antibiotic resistance genes in Chinese swine farms[J]. Proceedings of the National Academy of Sciences of the United States of America, 110(9): 3435–3440.
DOI:10.1073/pnas.1222743110
|
Zhu Y G, Zhao Y, Li Bing, et al. 2017. Continental-scale pollution of estuaries with antibiotic resistance genes[J]. Nature Microbiology, 2: 1–7.
|
Zhang X X, Zhang T, Fang H. 2009. Antibiotic resistance genes in water environment[J]. Applied Microbiology and Biotechnology, 82(3): 397–414.
DOI:10.1007/s00253-008-1829-z
|
Zhang X, Zhang T. 2011. Occurrence, abundance, and diversity of tetracycline resistance genes in 15 sewage treatment plants across China and other global locations[J]. Environmental Science & Technology, 45: 2598–2604.
|