 2017, Vol. 37
2017, Vol. 37



2. 四川职业技术学院, 遂宁 629000
2. Sichuan Vocational and Technical College, Suining 629000
矿产资源的开采、冶炼及含砷和其它金属制品的使用导致周围环境中砷与其它重金属的复合污染,如多种木材防腐剂CCA(Chromated-Copper-Asenate)的使用带来环境As-Zn的复合污染(Choi et al., 2012),含砷农药的长期使用造成As-Pb元素在土壤中的累积(王海慧等,2009).重金属污染不仅破坏土壤生态环境,导致生物多样性下降,而且还威胁人类健康,因此,治理土壤重金属污染成为人们的共识.常用的土壤重金属污染治理修复技术有以下几种(罗磊等,2008;叶文玲等,2014):固定、稳定化技术、土壤淋洗技术、原位电动修复技术和生物修复技术等,但这些方法的治理成本比较高,并且对环境扰动较大.与以上方法相比,植物修复技术无二次污染、操作容易、成本费用更低(李有志等,2012).
近年来,关于土壤重金属污染的植物修复技术的研究也越来越多,而超富集植物是植物修复的基础.超富集植物是一种可在植物体内超量积累重金属并能将其大量转运到地上部分的特殊植物(王英辉等,2007).蜈蚣草是世界上首次报道发现的砷超富集植物,同时也是我国境内发现的第一种重金属超富集植物(谢景千等,2010).蜈蚣草的耐砷特性和对砷的富集能力极强,它的最大含砷量比普通植物高数万倍、甚至数十万倍(李小林等,2011).除砷以外,蜈蚣草对铅、锌等重金属也具有一定的富集能力,在重金属污染土壤的生物修复中具有很好的应用价值和潜力(刘月莉等,2009).共附生细菌是指与动植物有关的细菌,包括附着在动植物表面的细菌及生长在其内部并与动植物存在着互惠互利或一方受益或寄生等关系的细菌.在重金属污染环境中,植物共附生细菌的种群结构、生理代谢会产生各种变化以响应重金属的胁迫.共附生菌还可以通过对重金属的吸附富集、氧化还原、成矿沉淀、淋滤、协同植物吸收等作用修复环境重金属污染(周耀渝等,2012;谢学辉等,2012).研究超富集植物共附生细菌的种群结构对于发现有用微生物资源并应用于土壤重金属污染的治理具有重要意义.
为探索铅锌矿区蜈蚣草共附生细菌在砷胁迫下的群落结构变化特征,本文利用变性梯度凝胶电泳(DGGE)技术,对四川省汉源县万顺铅锌矿区蜈蚣草根状茎中共附生细菌多样性和群落结构进行研究,并分析共附生细菌群落结构与蜈蚣草根状茎中有效重金属(Cu、Cd、Pb、Zn和As)间的关系.以期挖掘重金属污染地区植物共附生菌资源,探究蜈蚣草共附生细菌的生物修复潜能,为深入认识重金属污染环境下共附生细菌与植物间的互作机制,提高土壤重金属污染生态修复的效率提供基础依据.
2 材料与方法(Materials and methods) 2.1 样品采集汉源县万顺铅锌矿区(29°23′17″N,102°38′02″E)位于四川省流沙河下游左岸,海拔约800~1200 m,区域四季分明,年平均气温17.9 ℃,年均降雨量780 mm,无霜期300 d,野外调查发现,蜈蚣草在该矿区广泛分布.根据采矿、冶炼、废渣排放等过程中产生的不同污染程度的土壤重金属污染矿区情况,在距尾矿堆积区不同距离处采样.共分为5个采样区,每个采样区重复采样3次.5个采样区分别为尾矿区、弃渣场、矿口和选矿区4个受铅锌矿区污染影响的区域及1个远离矿山的农田区(对照),编号依次为WK、QZ、KK、XK和CK.随机采集多株生长良好的蜈蚣草,装入无菌密封塑料袋中,贴上标签,立即带回实验室,置于4 ℃冰箱保存.
2.2 蜈蚣草根状茎重金属含量分析将蜈蚣草地下部根状茎用自来水冲洗干净,再用去离子水淋洗2~3次,于105 ℃下杀青30 min,在80 ℃下烘干至恒重,磨碎后过60目尼龙筛,用HNO3-HC1O4消解,采用AFS-230E型原子荧光光度计测定Cu、Cd、Zn、Pb、As的有效含量(郑有坤等,2014).样品分析过程中采用国家标准参比物质(植物:GBW-08501) 进行分析质量控制.
2.3 蜈蚣草共附生细菌群落结构分析 2.3.1 蜈蚣草根状茎总DNA的提取将打碎的蜈蚣草根状茎样品置于无菌研钵中,加液氮、磷酸缓冲液,研磨,磨细后装入2 mL无菌EP管中,再加1.5 mL磷酸缓冲液,配平(Tian et al., 2007);然后加10 μL蛋白酶K,放入37 ℃培养箱,放置30 min,隔10 min摇1次;再加入200 μL 20%SDS(十二烷基硫酸钠),65 ℃水浴2 h,隔20 min摇1次;在离心机上以6000 r · min-1离心10 min,将上清液转入新的离心管中.在原管中重新加入500 μL提取液和25 μL 20%SDS,用无菌牙签混匀后,在65 ℃水浴锅中水浴10 min,再在离心机上以6000 r · min-1离心10 min,取上清液合并.在上清液中加入0.5倍体积的50% PEG,在4 ℃下静置过夜;然后于4 ℃、6000 r · min-1的条件下离心10 min,取上清液合并.用1倍体积的苯酚、氯仿、异戊醇抽提3次,抽提时使用的离心机设置为10000 r · min-1,离心15 min.加入0.1倍体积的3 mol · L-1的NaAc和0.6倍体积的异丙醇沉淀1 h,再在离心机上以1000 r · min-1离心25 min.加入预冷的70%乙醇5 μL进行洗涤,10000 r · min-1离心25 min后弃上清液.待乙醇完全挥发后,加入100 μL的1×TE溶解,保存.
2.3.2 16SrRNA基因扩增采用巢氏PCR对目标进行扩增(张波等,2013),以8-27F和1523-1504R引物进行第一轮扩增,引物序列分别为8-27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和1523-1504R(5’-AAGGAGGTGATCCAGCCGCA-3’).PCR反应体系为20 μL,包括:10 μL mix,0.2 μL 8-27F引物,0.2 μL 1523-1504R引物,1 μL DNA模板和8.6 μL无菌水.PCR反应条件为:94 ℃预变性5 min;94 ℃变性1 min,56 ℃退火1 min,72 ℃延伸2 min,30个循环;72 ℃最终延伸10 min.
第二轮扩增以第一轮的扩增产物为模板,以F341-GC和R518为引物进行PCR扩增.其中,引物序列为F341-GC(5’-CGCCCG GCG CGGGGG TCC GGGAGGCAGCAG-3’)和R518(5’-ATT ACC GCG GCT GG-3’).PCR反应体系为50 μL,包括:25 μL mix,0.5 μL F341-GC引物,0.5 μL R518引物,1 μL DNA模板和23 μL无菌水.PCR反应条件为:95 ℃预变性6 min;94 ℃变性1 min,65 ℃退火45 s,72 ℃延伸45 s,每个循环退火温度降低0.5 ℃,直至55 ℃,20个循环;然后94 ℃变性1 min,55 ℃退火45 s,72 ℃延伸45 s,15个循环;72 ℃最终延伸10 min.产物于1.5%琼脂糖凝胶电泳检测后用上海生工的SanPrep柱式PCR产物纯化试剂盒对PCR产物进行纯化.
2.3.3 变性梯度凝胶电泳在Dcode系统(Bio-Rad公司)上进行DGGE分析,聚丙烯酰胺凝胶浓度为8%,变性剂浓度范围为30%~50%,在1×TAE缓冲液中,以120 V电压、60 ℃恒温电泳5~6 h(邢薇等,2008).共获得32个有效序列,在NCBI中的序列登录号为KU844118~KU844149.
2.4 数据处理与分析基础数据的处理利用Excel 2010进行,Duncan单因素方差分析用SPSS17.0完成.DGGE图谱利用Bio-Rad公司的Quantity One 4.4分析.用MEGA 5.0,采用Neighbor-joining法(Tamura et al., 2011)构建系统发育树.用Shannon多样性指数(H)、丰富度指数(S)和均匀度指数(E)等评价群落结构多样性(沈烁等,2015).采用CANOCO4.5软件(Microcomputer Power Ithaca USA)分析群落结构和蜈蚣草重金属含量间的关系.多样性指数计算公式如下:
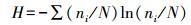
|
(1) |
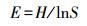
|
(2) |
式中,ni为单一条带的强度,N为所有条带的总强度,S为每一泳道总的条带数.
3 结果(Results) 3.1 蜈蚣草重金属含量分析对蜈蚣草根状茎内部的5种重金属(Cu、Cd、Zn、Pb和As)有效含量进行测定,表 1结果显示,不同采样点蜈蚣草根状茎中的Cu、Cd、Zn、Pb和As 5种重金属的有效含量存在明显差异(p < 0.05).Cu、Cd、Pb和As 4种重金属均以采自矿口区的样品含量最高,而Zn则以采自尾矿区的样品含量最高,重金属含量最低的样品是采自农田区域的样品.矿区蜈蚣草中有效Cu的含量是农田区域蜈蚣草中的6倍,有效Cd含量是农田区域蜈蚣草中的4倍,有效Zn、Pb含量均是农田蜈蚣草中的6倍,有效As含量也高出农田蜈蚣草中近3倍,显示矿区蜈蚣草对本试验测定的5种重金属均表现出明显的富集现象.
| 表 1 不同采样点蜈蚣草中有效重金属含量 Table 1 The available heavy metals contents of Pteris vittata in different sampling sites |
汉源铅锌矿区蜈蚣草根状茎共附生细菌的种群结构DGGE分析见图 1.蜈蚣草共附生细菌DGGE的图谱显示,不同的特异性条带代表着不同的细菌种群.从DGGE图谱可以看出,每个样品都呈现出数目不等的条带,但各个泳道条带数量明显不同,且每个泳道条带的颜色也都不相同,说明铅锌矿区不同程度重金属污染土壤上生长的蜈蚣草根状茎共附生细菌种群存在着明显差异.对比不同样品,以矿口和尾矿区样品的电泳条带数目最多、最复杂,且条带亮度最高.除弃渣区外,农田蜈蚣草样品的条带数目最少、最简单,亮度也较低.各泳道也有共同的条带,表明各采样点的蜈蚣草根状茎存在相同的共附生菌群,如WK1-1、WK1-2、KK3-2和XK1-4为所有泳道中共有的条带,显示这些内生细菌是铅锌矿区蜈蚣草根状茎的优势共有菌群.
 |
| 图 1 铅锌矿区不同区域蜈蚣草共附生细菌DGGE图谱 Fig. 1 DGGE analysis of the symbiotic or epiphytic bacteria in P. vittata in different regions of Pb-Zn mining |
基于DGGE图谱对蜈蚣草根状茎共附生细菌多样性指数(H)、丰富度指数(S)和均匀度指数(E)进行计算,结果见表 2.可以看出,不同位置采集的蜈蚣草样品共附生细菌多样性指数、丰富度指数及均匀度指数都存在差异.蜈蚣草根状茎共附生细菌群落结构的多样性指数和丰富度指数有高度一致性,皆表现为尾矿>矿口>弃渣区>选矿区>农田.总体而言,多样性指数为2.39~3.29,丰富度指数为11~27,均匀度指数为0.997~0.998.其中,尾矿区域蜈蚣草的共附生细菌的多样性指数和丰富度指数最高,分别达到了3.29和27,多样性指数和丰富度指数最低的是农田区,分别为2.39和11, 而在均匀度部分差异不显著.
| 表 2 蜈蚣草共附生细菌的多样性指数、丰富度指数、均匀度指数 Table 2 Index of the Shannon-Wiener diversity(H), evenness(E), and richness(S) of the symbiotic or epiphytic bacteria in Pteris vittata |
对蜈蚣草样品各重金属含量与多样性指数、丰富度指数和均匀度指数进行相关性分析,结果见表 3.可见,样品多样性指数和丰富度指数都与Cu、Cd、Pb、As呈显著正相关(p < 0.01),与Zn的相关系数为正,但未达到显著水平.均匀度指数与Cu、Cd含量显著正相关(p < 0.05),与Pb和As显著正相关(p < 0.01),与Zn的相关系数为正,但未达到显著水平.其中,As和多样性指数(H)间的相关系数最高,表明As含量对细菌多样性影响较大.
| 表 3 蜈蚣草重金属含量与共附生细菌多样性指数的相关性分析 Table 3 Correlation analysis between the heavy mental amounts and Shannon-Wiener diversity index(H), evenness(E), and richness(S) of symbiotic or epiphytic bacteria in Pteris vittata |
根据DGGE图谱,选取特异性和共有条带进行切胶回收克隆测序,共得到32条有效序列.通过Genbank数据库中Blast比对,选取与目标同源性最大的16S rRNA序列作为参比序列共同构建系统发育树,结果如图 2所示.通过分析系统发育树可知,大部分序列隶属于变形菌门(Proteobacteria)和厚壁菌门(Firmicutes),还有少量属于放线菌门(Actinobacteria),说明铅锌矿区蜈蚣草共附生细菌较为丰富.在16S rRNA的比对分析中,所有的样品序列和GenBank中已知序列的相似性在98%~100%之间.菌群结构可分为3部分:第1部分包含23个克隆条带,全部隶属于变形菌门(Proteobacteria),其中,10个序列属于α-变形菌(α-Proteobacteria),包括弯曲杆菌属(Campylobacter)和根瘤菌属(Rhizobium);8个序列属于β-变形菌(β-Proteobacteria),主要属于伯克氏菌属(Burkholderia);5个序列属于γ-变形菌(γ-Proteobacteria),包括假单胞菌属(Pseudomonas)和肠杆菌属(Enterobacteria)等类群.第2部分包含8个克隆条带,全部属于厚壁菌门(Firmicutes)的芽孢菌属(Bacillus).KK1-1单独与其参比菌株成构成第3部分,与链霉菌属(Streptomyces)的相似度达到99%.上述结果表明,在蜈蚣草根部共附生细菌中α-变形菌(α-Proteobacteria)是优势菌群.结合DGGE图谱,条带WK1-1、KK3-2和XK1-4为5个样品共有,分别与根瘤菌属(Rhizobium)、伯克氏菌属(Burkholderia)、弯曲杆菌属(Campylobacter)相似.矿口区域所含的细菌类型丰富,大部分属于变形菌门
 |
| 图 2 基于DGGE回收条带的细菌16S rRNA基因系统发育树 Fig. 2 Phylogenetic analysis of 16S rRNA gene sequences from DGGE bands |
(Proteobacteria)和厚壁菌门(Firmicutes).KK1-1是矿口区、选矿区和尾矿区的共有优势条带,属于放线菌中的链霉菌属(Streptomycete).尾矿区域的共附生细菌均属于变形菌门(Proteobacteria),其中与根瘤菌属(Rhizobium)、假单胞菌属(Pseudomonas)、伯克氏菌属(Burkholderia)的相似度最高.QZ2-1为弃渣区特有的条带,属于根瘤菌属(Rhizobium).XK1-2和XK1-3也仅在选矿区域检测到,二者都属于芽孢杆菌属(Bacillus).作为对照样的农田样品优势共附生细菌包括短波单胞菌属(Brevundimonas)、极小单孢菌属(Pusillimonas)和芽孢杆菌属(Bacillus)等.上述结果显示,铅锌矿区蜈蚣草存在丰富的共附生细菌种群.
3.6 蜈蚣草共附生细菌群落与根状茎重金属含量间的关系运用Canoco4.5软件,根据测得的5个点共15个样品的蜈蚣草重金属含量和共附生细菌群落多样性、丰富度、均匀度指数进行RDA分析,探究细菌群落在重金属胁迫下的变化.RDA分析结果如图 3所示,第一排序轴特征值较高,解释了影响共附生细菌结构因素的87.5%,第一轴和第二轴的物种-环境相关性系数分别为0.939和0.610,共附生细菌群落结构及土壤重金属含量相关系数均为1,表明排序能够很好地反映共附生细菌与重金属含量之间的关系.
 |
| 图 3 蜈蚣草共附生细菌群落与重金属含量的冗余分析 Fig. 3 The redundant analysis (RDA) of symbiotic or epiphytic bacterial community and heavy metal contents of P. vittata |
在共附生细菌与重金属含量的相关系数中,第一轴与As、Pb、Cu、Cd的相关性较高,相关系数分别为0.8956、0.8808、0.7335、0.6753,第二轴与Zn的相关性较高,相关系数为0.4785(图 3).环境变量箭头越长,对物种变量的影响越大.由图可知,Cd对细菌群落的影响相对较小,而其它重金属的影响相对较高,显示铅锌矿区蜈蚣草根状茎细菌群落结构与这些重金属具有相关性.RDA排序图中,弃渣区、尾矿区和矿口区的共附生细菌与重金属含量呈正相关.对于农田和选矿区的细菌群落,则与重金属呈负相关,重金属含量提高会抑制蜈蚣草共附生细菌群落结构.
4 讨论(Discussion)物种多样性是指细菌物种的复杂程度,是衡量群落规模和重要性的基础(林海等,2014).本研究中不同采样点蜈蚣草共附生细菌多样性指数、丰富度指数及均匀度指数都存在一定差异.尾矿和矿口两处重金属含量最高的区域,多样性指数和丰富度指数最高,而重金属含量最低的农田区域多样性指数和丰富度指数最低.结合蜈蚣草重金属含量和与共附生细菌多样性指数的相关性分析,样品多样性指数和丰富度指数与Cu、Cd、Pb、As呈显著正相关(p < 0.01),这与相关报道相似,如湛方栋等(2007)对铅锌矿区和非矿区毛兽蝇子草根际细菌群落结构的研究发现,矿区毛萼蝇子草根际耐Pb细菌和耐Cd细菌的数量均极显著高于非矿区,对Pb,Cd的耐受程度和种群多样性也明显高于非矿区.王义聪(2014)在对云南铅锌矿区的根际细菌种群结构多样性研究发现,受重金属胁迫下,紫茎泽兰共附生菌群落结构高于非矿区.这说明长期重金属胁迫导致植物共附生细菌群落结构发生明显变化.
本研究中,重金属对蜈蚣草共附生细菌群落结构的影响并不是简单的线性关系.冗余分析(RDA)显示,在重金属含量较高的采样点,重金属对蜈蚣草共附生细菌群落结构有促进作用,在重金属含量相对较低的农田区域,蜈蚣草共附生细菌群落与砷呈负相关性.相关研究也表明,As对蜈蚣草的影响十分复杂,在轻度、中度As污染区中,蜈蚣草地上部As含量随着根际土壤As含量的增加而增加,在重度As污染区中,蜈蚣草地上部的As含量却随根际土壤As含量的增加逐渐降低(申红玲等,2014).在低As处理下,蜈蚣草植株有抑制As进入蜈蚣草细胞的趋势.随着As含量的提高,蜈蚣草对As的累积量有明显增加(肖细元等,2008).此外,对于其它重金属的污染,蜈蚣草的生长也存在着不同的响应.相关研究发现,在栽培蜈蚣草的土壤中添加不同剂量重金属Pb、Cu、Zn后,会导致蜈蚣草生长量出现明显的变化,但变化趋势不尽一致.随Pb、Zn添加量的增加,蜈蚣草生物量也呈不同程度的增加, 但添加量非常高时,Pb和Zn对蜈蚣草生长均有一定的抑制作用(潘志明等,2007).
群落结构分析表明,蜈蚣草共附生细菌种群结构多样性丰富,包括变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和放线菌门(Actinobacteria),其中,变形菌门细菌是蜈蚣草根状茎的优势共附生细菌,占总分离序列的71%,主要包括伯克氏菌属(Burkholderia)、肠杆菌属(Enterobacteria)、假单胞菌属(Pseudomonas)和根瘤菌属(Rhizobium)等.这与之前对其它重金属矿区植物共附生细菌的研究结果相似,如对铅锌矿区东南景天植物共附生细菌群落进行分析发现,变形菌为主要的优势菌种(Chen et al., 2014);对紫金泽兰根际细菌群落结构的研究中也发现了类似的结果(牛红榜等,2007).在植物根部重金属含量较高的矿口和尾矿区,细菌群落主要属于芽孢杆菌属(Bacillus)、链霉菌属(Streptomyces)、伯克氏菌属(Burkholderia)、根瘤菌属(Rhizobium)和假单胞菌属(Pseudomonas).对于重金属含量较低的农田区域,细菌群落主要为根瘤菌属(Rhizobium)和芽孢杆菌属(Bacillus).相关微生物强化蜈蚣草富集砷能力的研究发现,将从砷污染土壤中提取的链霉菌Streptomyces lividans PSQ加入到蜈蚣草的培养中,可以显著提高蜈蚣草对砷污染土壤的修复效率(赵根成等,2010).而在其它的修复砷污染土壤的研究中,对于蜈蚣草,施用砷酸还原菌明显促进了植物地上部对砷的吸收(增长了13%~110%)和生长(增长了93%~290%),同时将植物地上部对砷污染土壤的修复效率提高了67%~478%(吴佳等,2011).当前治理砷污染所用的微生物菌株多属于链霉菌属(Streptomyces)、芽孢杆菌属(Bacillus)(纪冬丽等,2016)、丛毛单胞菌属(Comamonas)(段志斌等,2016).对于铅锌矿区蜈蚣草共附生细菌的研究发现类似的现象,如在矿口发现的KK-1为链霉菌属(Streptomyces);KK1-10、KK3-6及选矿区的XK1-2、XK1-3等均与芽孢杆菌(Bacillus)有着99%以上的相似度;但对于丛毛单胞菌属(Comamonas),在本研究中并未发现.而对于从所有样品中均检测到的优势菌——根瘤菌属(Rhizobium)和伯克氏菌属(Burkholderia),未见用于治理土壤砷污染的相关报道.除了用于治理砷污染以外,从蜈蚣草中分离到的细菌还被报道用于治理其它重金属污染,如具有活化重金属和多种促生特性的Burkholderia sp. J62显著增加了玉米和番茄的生物量及其根际醋酸铵提取态的Pb和Cd,促进了植物对重金属的吸收(Jiang et al., 2008).菌株Pseudomonas sp. RJIO和Bacillus sp. RJ16能够活化土壤中的Pb和Cd,且具有多种促生特性,将这些菌株接种番茄后可以有效地促进土壤Pb和Cd的去除(He et al., 2009).综上,蜈蚣草根状茎的共附生细菌种群在重金属污染土壤的治理方面具有潜在应用价值.
5 结论(Conclusions)以免培养手段分析了铅锌矿区蜈蚣草根状茎共附生细菌群落结构对重金属的响应特征,结果发现,不同采样点蜈蚣草共附生细菌的群落结构和多样性存在一定的差异,多样性指数和丰富度指数表现为尾矿>矿口>弃渣区>选矿区>农田.细菌多样性指数和丰富度指数都与Cu、Cd、Pb、As呈显著正相关(p < 0.01),与Zn的相关系数为正,但相关性不显著,其中,As对细菌多样性影响最大.伯克氏菌属(Burkholderia)、肠杆菌属(Enterobacteria)、假单胞菌属(Pseudomonas)、根瘤菌属(Rhizobium)、弯曲杆菌属(Campylobacter)和链霉菌属(Streptomyces)等是蜈蚣草的优势共附生细菌种群.
Chen B, Zhang Y, Rafiq M T, et al. 2014. Improvement of cadmium uptake and accumulation in Sedum alfredii by endophytic bacteria Sphingomonas SaMR12:Effects on plant growth and root exudates[J]. Chemosphere, 117: 367–373.
DOI:10.1016/j.chemosphere.2014.07.078
|
Choi Y S, Ahn B J, Kim G H. 2012. Extraction of chromium, copper, and arsenic from CCA-treated wood by using wood vinegar[J]. Bioresource Technology, 120: 328–331.
DOI:10.1016/j.biortech.2012.06.045
|
陈同斌, 阎秀兰, 肖细元, 等. 2008. 磷肥对砷超富集植物蜈蚣草修复砷污染土壤的影响[J]. 环境科学, 2008, 29(10): 2906–2911.
DOI:10.3321/j.issn:0250-3301.2008.10.038 |
段志斌, 胡丰青, 安吉平, 等. 2016. 砷污染及其植物修复技术进展[J]. 现代农业科技, 2016(14): 190–193.
DOI:10.3969/j.issn.1007-5739.2016.14.112 |
He L Y, Chen Z J, Ren G D. 2009. Increased cadmium and lead uptake of a cadmium hyperaccumulator tomato by cadmium-resistant bacteria[J]. Ecotoxicology and Environmental Safety, 72(5): 1343–1348.
DOI:10.1016/j.ecoenv.2009.03.006
|
Jiang C Y, Heng X F, Qian M. 2008. Isolation and characterization of a heavy metal-resistant Burkholderia sp.from heavy metal-contaminated paddy field soil and its potential in promoting plant growth and heavy metal accumulation in metal-polluted soil[J]. Chemosphere, 72(2): 157–164.
DOI:10.1016/j.chemosphere.2008.02.006
|
纪冬丽, 孟凡生, 薛浩, 等. 2016. 国内外土壤砷污染及其修复技术现状与展望[J]. 环境工程技术学报, 2016, 6(1): 90–99.
|
李小林, 颜森, 张小平, 等. 2011. Pb-Zn矿区重金属污染对微生物数量及放线菌群落结构的影响[J]. 农业环境科学学报, 2011, 30(3): 486–475.
|
李有志, 罗佳, 张灿明, 等. 2012. 湘潭锰矿区植物资源调查及超富集植物筛选[J]. 生态学杂志, 2012, 31(1): 16–22.
|
林海, 崔轩, 董颖博, 等. 2014. 铜尾矿重金属Cu、Zn对细菌结构群落的影响[J]. 中国环境科学, 2014, 34(12): 3182–3188.
|
刘月莉, 伍钧, 唐亚, 等. 2009. 四川甘洛铅锌矿区优势植物的重金属含量[J]. 生态学报, 2009, 29(4): 2020–2026.
|
罗磊, 张淑贞, 马义兵. 2008. 土壤中砷吸附机理及其影响因素研究进展[J]. 土壤, 2008, 40(3): 351–359.
|
牛红榜, 刘万学, 方万浩, 等. 2007. 紫茎泽兰根际土壤中优势细菌的筛选鉴定及拮抗性能评价[J]. 应用生态学报, 2007, 18(12): 2795–2800.
|
潘志明, 邓天龙. 2007. 砷污染土壤的蜈蚣草修复研究[J]. 土壤, 2007, 39(3): 341–346.
|
申红玲, 何振艳, 麻密. 2014. 蜈蚣草砷超富集机制及其在砷污染修复中的应用[J]. 植物生理学报, 2014, 50(5): 591–598.
|
沈烁, 杨长明, 成水平. 2015. PCR-DGGE分析合肥市塘西河表层沉积物细菌群落结构空间分布特征[J]. 应用与环境生物学报, 2015, 21(1): 80–87.
|
Tamura K, Peterson D, Peterson N, et al. 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Molecular Biology and Evolution, 28(10): 2731–2738.
DOI:10.1093/molbev/msr121
|
Tian X L, Cao L X, Tan H M, et al. 2007. Diversity of cultivated and uncultivated actinobacterial endophytes in the stems and roots of rice[J]. Microbial Ecology, 53(4): 700–707.
DOI:10.1007/s00248-006-9163-4
|
王海慧, 郇恒福, 罗瑛, 等. 2009. 土壤重金属污染及植物修复技术[J]. 中国农学通报, 2009, 25(11): 210–214.
|
王义聪. 2014. 云南募乃露天铅锌矿区优势植物根内生细菌分子多样性研究[D]. 昆明: 云南大学
http://cdmd.cnki.com.cn/Article/CDMD-10673-1014339925.htm |
王英辉, 陈学军, 祁士华. 2007. 铅污染土壤的植物修复治理技术[J]. 土壤通报, 2007, 8(4): 790–794.
|
韦朝阳, 郑欢, 孙歆, 等. 2008. 不同来源蜈蚣草吸收富集砷的特征及植物修复效率的探讨[J]. 土壤, 2008, 40(3): 474–478.
|
吴佳, 谢明吉, 杨倩, 等. 2011. 砷污染微生物修复的进展研究[J]. 环境科学, 2011, 32(3): 817–824.
|
谢景千, 雷梅, 陈同斌, 等. 2010. 蜈蚣草对污染土壤中As、Pb、Zn、Cu的原位去除效果[J]. 环境科学学报, 2010, 30(1): 165–171.
|
谢学辉, 范风霞, 袁学武, 等. 2012. 德兴铜矿尾矿重金属污染对土壤中微生物多样性的影响[J]. 微生物学通报, 2012, 39(5): 624–637.
|
邢薇, 左剑恶, 林甲, 等. 2008. 20℃ EGSB反应器中颗粒污泥的微生物种群结构分析[J]. 环境科学, 2008, 29(9): 2558–2563.
|
叶文玲, 樊霆, 鲁洪娟, 等. 2014. 蜈蚣草的植物修复作用对土壤中砷总量及形态分布的影响研究[J]. 土壤通报, 2014, 45(4): 1003–1007.
|
湛方栋, 何永美, 祖艳群, 等. 2007. 铅锌矿区和非矿区毛萼蝇子草(Silene pubicalycina)根际细菌群落结构研究[J]. 农业环境科学学报, 2007, 26(s2): 524–528.
|
张波, 李小林, 张小平, 等. 2013. 花首稽内生放线菌多样性及群落结构[J]. 草业学报, 2013, 22(5): 113–114.
DOI:10.11686/cyxb20130513 |
赵根成, 廖晓勇, 阎秀兰, 等. 2010. 微生物强化蜈蚣草累积土壤砷能力的研究[J]. 环境科学, 2010, 31(2): 431–436.
|
郑有坤, 任丹, 卢圣鄂, 等. 2014. 铅锌矿区蜈蚣草内生放线菌的多样性及群落结构[J]. 环境科学学报, 2014, 34(11): 2832–2838.
|
周耀渝, 杨胜香, 袁志忠, 等. 2012. 湘西铅锌矿区重金属污染评价及优势植物重金属累积特征[J]. 地球与环境, 2012, 40(3): 361–366.
|