 2017, Vol. 37
2017, Vol. 37



铀是核电产业和核技术应用领域中非常重要的战略性资源,随着铀矿的大量开采和湿法冶金技术的成熟,产生了大量的浓度低于1 mg·L-1的含铀废水,这些低浓度的含铀废水随着环境的迁移和食物链的富集,不可避免地会进入环境,从而对自然生态环境和人类健康构成潜在的威胁(代群威等,2012; Das,2012;Ding et al.,2014).近年来,国内外众多研究人员广泛关注于高浓度含铀废水(初始铀浓度大于50 mg·L-1)的处理,并取得了显著的成果(邓钦文等,2014;彭国文等,2012;夏良树等,2008;Pang et al.,2011),然而对于低浓度的含铀废液(放射性活度浓度范围为37~3.7×105 Bq·L-1(魏广芝等,2007))的处理关注却相对较少(Ding et al.,2014;Li et al.,2014;Xiao et al.,2015),且浓度越低处理难度越大.目前,化学(混凝)沉淀、离子交换、蒸发浓缩、吸附是处理放射性含铀废水的基本工艺,但在实际应用中这些方法对于处理离子浓度在1~100 mg·L-1的低浓度含铀废水,存在着高能耗、低效率、高成本、需二次废物处理等问题(林莹等,2009;Wang et al.,2009; Sarada et al.,2014).多年来,研究人员一直致力于探寻新的高效经济的低浓度含铀废水的处理技术,1991年美国学者首次揭示了微生物能够还原铀,开创了微生物与铀相互作用的新时代,微生物处理含铀废水的技术从而得到了广泛的重视和发展(胡恋等,2007).同时,研究人员们还发现,这些生物吸附剂拥有良好的金属结合能力,在特殊条件下还能实现特异性选择吸收某些金属离子,能够很好地降低溶液中金属离子浓度(从μg·L-1降低到ng·L-1)(Wang et al.,2009; Fomina et al.,2014;陈灿等,2006).利用微生物吸附低浓度放射性废水是一种新型高效的处理方法,具有低成本、耗能少、无二次污染等优点,并能与其他工艺相结合实现工业生产化(Vijayaraghavan et al.,2008).
在众多微生物吸附剂(细菌、真菌、藻类)中,选用发酵工业的废弃菌体作为吸附剂,不仅可以降低生物吸附剂的成本,还可使其具有工业化应用的潜力.面包酵母作为一类重要且廉价的工业微生物,突显其作为生物吸附剂的优越性:安全无毒性、来源稳定易培养、理想模式生物可在分子水平上研究其吸附机理(陈灿等,2006).同时,研究发现,酵母菌可以去除多种重金属,并且对于放射性核素铀的吸附量较高,在竞争吸附中占据优势地位,受因素干扰较小(陈灿等,2006;Wang et al.,2006).目前,关于酵母菌对放射性核素铀的吸附能力和机理研究中,大部分研究者使用的都是活体酵母菌(Liu et al.,2010;Popa et al.,2003; Faghihian et al.,2012),也有部分研究人员研究死体酵母菌,发现其具有与活体酵母相似甚至更好的吸附效果,而对于活、死菌吸附机理的研究却很少有人关注(Lu et al.,2013).
本文基于近年来国内外的研究成果及自身的研究工作,探究活性与灭活面包酵母对低浓度放射性核素铀在不同酸碱度、吸附时间、吸附剂浓度条件下的吸附差异,并对吸附动力学特性进行分析,最后讨论其不同的吸附机理,以期为后续发展的生物吸附工艺提供一定的参考.
2 材料和方法(Materials and methods) 2.1 实验材料 2.1.1 主要仪器与试剂WGJ-Ⅲ型微量铀分析仪,恒温培养摇床,TG16-WS台式高速离心机,立式压力蒸汽灭菌器,傅立叶变换-红外光谱仪,JSM-6701F冷场发射型扫描电镜.硝酸铀酰及其他试剂均为分析纯.
2.1.2 菌株的选择与制备实验所用菌种为市售的安琪酵母有限公司生产的面包酵母.称取一定量的干酵母加入到2%的灭菌葡萄糖溶液中活化,将活化后的菌体种子液接入到无菌YPD(1%酵母提取物、2%蛋白胨、2%葡萄糖)液体培养基中,于恒温培养摇床(30 ℃,200 r·min-1)中振荡培养30 h后,离心(5000 r·min-1,5 min)收集菌体,将收集到的菌体用无菌去离子水反复清洗3次,然后将其中一部分稀释成菌体悬浮液,置于4 ℃ 冰箱中保存备用;将另一部分稀释的菌体悬浮液在0.15 MPa、121 ℃ 立式压力蒸汽灭菌器中灭活20 min,得到灭活酵母,置于4 ℃ 冰箱中保存备用.
2.1.3 铀溶液配置准确称取一定量的硝酸铀酰[UO2(NO3)2·6H2O],溶解于0.01 mol·L-1的硝酸溶液中,配制成1 mg·mL-1的标准储备铀溶液,然后用0.001 mol·L-1硝酸水逐级稀释得到10 mg·L-1的实验用铀溶液.实验过程中使用NaOH、HNO3调节溶液pH值,为了减小外加阳离子对吸附过程的影响,在批次实验中Na+离子强度设置为0.01 mol·L-1.
2.2 实验方法 2.2.1 吸附实验取容积为40 mL的离心管,根据不同实验目的,加入一定体积的活性酵母菌悬浮液和3 mL 0.1 mol·L-1硝酸钠溶液于摇床上恒温振荡混合(200 r·min-1,30 min)后,加入3 mL 10 mg·L-1铀溶液(保持吸附系统中初始铀离子浓度为 1 mg·L-1),在摇床上恒温振荡吸附(200 r·min-1),一定时间后取样离心(10000 r·min-1,15 min),取上层清液测定其中铀离子浓度.灭活酵母菌与活性酵母菌实验操作过程完全一致.
2.2.2 计算方法铀浓度利用微量铀分析仪工作曲线法测定,铀的吸附量及吸附率用下式表示:

|
(1) |
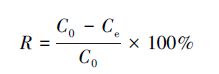
|
(2) |
式中,Q为吸附量(mg·g-1),R为吸附率,C0为初始溶液中铀离子浓度(mg·L-1),Ce为吸附后溶液中铀离子浓度(mg·L-1),Cm为加入酵母菌悬浮液的质量浓度(g·L-1)(代群威等,2008).
2.3 分析测试由于铀溶液浓度太低不利于样品的观测,因此,在制作分析样品时,将铀溶液浓度提高至100 mg·L-1.
2.3.1 扫描电镜样品制备离心收集吸附前后的活性酵母与灭活酵母菌体,加入2.5%戊二醛溶液固定1 h后除去戊二醛,清洗后用洁净的胶头滴管将菌体置于洁净抛光的锡箔纸上,依次用20%、50%、80%、100%的乙醇溶液梯度脱水处理,待自然晾干后,喷金处理,置于JSM-6701F冷场发射型扫描电镜下观测,得到菌体的显微图像.
2.3.2 红外光谱样品制备离心收集吸附前后的活性酵母与灭活酵母菌体,置于30 ℃恒温干燥箱中干燥备用.采用KBr压片法,在相同的频率波长下测定红外吸收光谱.
3 结果与分析(Results and analysis) 3.1 pH值对铀吸附效果的影响吸附系统pH值不仅决定着酵母菌表面电荷位点分布和硝酸铀酰在液相中的水解及其络合形态,还会影响生物吸附剂表面官能团的结构与性能(吴海云等,2010;Popa et al.,2003),因此,在实验中系统的酸碱度会影响铀离子与酵母菌表面吸附位点的结合程度.不同pH值下,活性酵母菌与灭活酵母对铀酰离子的吸附效果如图 1所示.
 |
| 图 1 pH值对不同活性酵母菌吸附铀的影响(a)及灭活酵母菌与活性酵母菌对铀吸附量比值(Q2/Q1)(b)(C0=1 mg·L-1,t=4 h,T=26 ℃,Cm=0.20 g·L-1) Fig. 1 Effect of pH on uranium adsorption by different active yeast(a)and the uranium adsorption ratio of dead yeast and live yeast(Q2/Q1)(b) |
从图 1a可以看出,pH值在3.0~5.0的范围内,活性酵母菌对铀的吸附率从8%增长到48%,基本呈线性增长趋势,灭活酵母菌对铀的吸附率始终保持在85%左右,变化不明显;而pH值在5.5~7.0的范围内,灭活酵母菌对铀的吸附率从84%下降至56%,呈线性下降趋势,活性酵母菌对铀的吸附率仅从50%下降至38%,下降趋势较缓.活性酵母菌对低浓度铀的最佳吸附pH值在5.5左右,吸附率达到50%,而灭活酵母菌的最佳吸附pH值在4.5左右,吸附率达到87%.从图 1b可以进一步看出,在酸性(pH=3)条件下,灭活酵母菌对铀的吸附量可以达到活性酵母菌的11倍,随着pH值的升高,灭活酵母菌与活性酵母菌的这种吸附差异逐渐减小,说明灭活酵母菌相对于活性酵母菌更能适应含低浓度铀的酸性水溶液.
3.2 时间对铀吸附效果的影响以时间为变量,研究了活性酵母与灭活酵母菌对铀离子吸附的动态平衡过程,结果如图 2所示.从图 2可以看出,吸附前30 min活性酵母菌与灭活酵母菌对铀的吸附速率近似相同,都保持较快的增长速率.此后,灭活酵母菌对铀的吸附速率趋于零,达到动态平衡状态,而活性酵母菌对铀的吸附速率趋缓,但仍保持着增长的趋势,240 min后才逐渐达到动态平衡,300 min后,灭活酵母菌对铀的吸附率(85%)明显高于活性酵母菌对铀的吸附率(70%).通常生物吸附过程被分为被动吸附和主动吸附两种模式,前者是一个快速过程,不需要消耗能量,主要依靠细胞壁上的官能团与金属离子之间的物理作用进行吸附,这个过程在活体与灭活的菌体吸附剂中都是主要过程;后者是一个相对缓慢的过程,需要依赖于能量及代谢系统的调控,主要通过细胞内酶促作用进行生物转运、生物沉淀和生物积累,仅见于活体吸附系统中(陈灿等,2006;Wang et al.,2006).因而对整个吸附过程来说,灭活酵母菌对铀的吸附是一个快速稳定的不消耗能量的过程,而活性酵母菌对铀的吸附则是一个相对缓慢的依靠自身代谢调控的过程.
 |
| 图 2 反应时间对不同活性酵母菌吸附铀的影响(pH=5.0±0.1,C0=1 mg·L-1,T=26 ℃,Cm=0.16 g·L-1) Fig. 2 Effect of time on uranium adsorption by different active yeast |
探究了活性酵母与灭活酵母初始菌浓度对铀的吸附率及吸附量的影响,结果如图 3所示.从图中 可以看出,当初始菌体投加量由0.064 g·L-1增至0.32 g·L-1时,活性酵母菌对铀的吸附率由62%提高到78%,灭活酵母菌仅从79%提高到87%.说明在低浓度铀的条件下,活性酵母菌对铀的吸附能力受初始菌浓度影响较大,随着菌浓度的升高,细胞表面吸附位点增加,有利于活性酵母菌对铀的吸附作用.当活性酵母菌与灭活酵母菌对铀的吸附系统到达相同状态时,活性酵母菌的初始浓度为0.32 g·L-1,而灭活酵母菌的初始浓度为0.064 g·L-1,仅为活性酵母菌浓度的1/5,且吸附量(12.42 mg·g-1)是活性酵母菌(2.45 mg·g-1)的6倍.进一步表明了在低浓度铀的条件下,灭活酵母菌对铀的吸附能力高于活性酵母菌,因此,在要求高效经济的吸附生产工艺中,应优先选用失活细胞体.
 |
| 图 3 不同活性菌体投加量对吸附铀的影响(pH=5.0±0.1,t=4 h,C0=1mg ·L-1,T=26 ℃;柱状图上方数据为吸附量) Fig. 3 Effect of biomass concentration on uranium adsorption by different active yeast |
在不同温度环境条件下,活性酵母和灭活酵母对铀的吸附能力如图 4所示.从图中可以看出,活性酵母菌(菌浓度0.32 g·L-1)对铀离子的最佳吸附温度为26 ℃,温度降低或升高,对于活性酵母菌吸附铀都有适当的抑制作用.这是由于温度较低时,酵母菌表面吸附位点的活性不强,而在较高温度下,这些表面吸附位点又失去了活性,从而说明活性酵母菌对铀的吸附过程要依赖于自身代谢系统的调控.而对于灭活酵母菌(菌浓度0.064 g·L-1),经过高温高压的作用,细胞表面的吸附位点已经失去活性,温度对灭活酵母菌的吸附能力影响不明显,但随着时间的延长,适当提高温度对吸附还是有促进作用,说明灭活酵母菌对铀的吸附是一种吸热反应,同时也表明灭活酵母菌是一种能够适应较宽温度范围的生物吸附剂(代群威等,2008).
 |
| 图 4 温度对不同活性酵母菌吸附铀的影响(pH=5.0±0.1,C0=1 mg·L-1) Fig. 4 Effect of temperature on uranium adsorption by different active yeast |
利用动力学分析可以进一步探讨酵母菌对铀酰离子的吸附机理,常用的离子动力学方程有准一级吸附动力学方程、准二级吸附动力学方程等.相比于准二级动力学模型,利用准一级动力学模型进行拟合需已知平衡吸附量,而对于生物吸附这种动态平衡过程来说,许多情况下平衡吸附量是未知的,因而用准二级动力学模型能够更好地揭示整个生物吸附的行为过程,并与速率控制步骤一致(Sarada et al.,2014).准二级动力学表达式如式(3)所示,经分离变量,积分可得式(4).
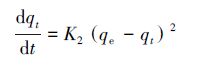
|
(3) |
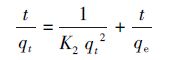
|
(4) |
式中,qe为平衡时的理论吸附量(mg·g-1),qt为t时刻的吸附量(mg·g-1),K2为准二级吸附速率常数(g·mg-1·min-1).
用式(4)分别对活性酵母与灭活酵母在不同温度下吸附铀的动力学过程进行研究,得到的拟合曲线如图 5所示,模型拟合参数见表 1.
 |
| 图 5 不同温度下不同活性酵母菌对铀的吸附动力学拟合(pH=5.0±0.1,C0=1 mg·L-1) Fig. 5 Kinetics of uranium adsorption by different active yeast at different temperature |
从表 1可以看出,不同温度下活性酵母菌与灭活酵母菌对铀离子的准二级动力学吸附速率方程拟合结果较好,R2均在0.99以上.一般来说,准二级动力学模型是假定吸附速率受化学吸附机理的控制,而这种化学吸附涉及到吸附剂与吸附质之间的电子共用或电子转移(Sarada et al.,2014),这就说明无论是活性酵母菌还是灭活酵母菌对铀的吸附过程中,都存在着化学吸附过程,并且这种化学吸附机制表现为菌体表面吸附位点与铀离子间的电子共用或电子转移.
| 表 1 不同温度下不同活性酵母菌对铀吸附的准二级动力学模型拟合参数 Table 1 Pseudo-second order adsorption kinetics parameters of uranium adsorption by different active yeast at different temperature |
图 6为吸附前后活性酵母与灭活酵母菌体的扫描电镜图,从图中可以明显看出,吸附铀前后活性酵母菌与灭活酵母菌菌体表面形貌的差异.吸附前活性酵母菌细胞表面完整光滑(图 6a),在与铀离子作用2 h后(图 6b),只有少量的铀离子沉淀物不均匀地附着在细胞表面,沉淀物聚集成小块状,同时细胞表面出现凹陷,形成空洞,表明铀离子开始与酵母细胞作用,这与刘明学等(2011)的研究结果一致.而经过高温高压灭活的酵母菌,在吸附铀离子前(图 6c),细胞虽保持完整,但表面出现褶皱,有明显突起,使其表面积增大,为铀离子的结合提供了更多的吸附位点,同时验证了灭活酵母菌的高吸附能力.由于灭活酵母菌对铀的吸附速率很快,仅30 min就达到动态平衡,因而在与铀离子作用2 h后(图 6d),大量的纳米颗粒状铀离子沉淀不均匀地附着在细胞表面,并成簇状向上生长.
 |
| 图 6 吸附前后活性与灭活酵母菌扫描电镜图(a.吸附前活性酵母菌,b.吸附后活性酵母菌,c.吸附前灭活酵母菌,d.吸附后灭活酵母菌;×15000;pH=5.0±0.1,t=2 h,C0=100 mg·L-1,Cm=1.60 g·L-1,T=30 ℃) Fig. 6 SEM images of different active yeast before and after adsorption of uranium |
图 7为吸附铀前后活性酵母菌与灭活酵母菌的红外光谱图.研究表明,吸附的铀酰离子主要沉积在细胞的表面即细胞壁上,因为细胞壁上的活性基团可以与金属离子发生定量的结合反应(白静等,2009).面包酵母菌细胞壁主要由葡聚糖、甘露聚糖、蛋白质和脂类,还有部分的几丁质(N-乙酰葡萄糖胺,以1-4葡萄糖苷键连接的多聚体)等组成(胡家骏等,1988).因面包酵母菌组成成分比较复杂,使得样品在整个波数段(400~4000 cm-1)内均有明显吸收,这样某些峰未能表现出来,一些不太灵敏的吸收峰被掩盖.
 |
| 图 7 吸附铀前后活性与灭活酵母菌红外光谱图(pH=5.0±0.1,t=2 h,C0=100mg·L-1,Cm=1.60 g·L-1,T=30 ℃) Fig. 7 FTIR results of different active yeast before and after adsorption of uranium |
从活性酵母菌吸附前后的红外光谱图(图 7)可见,活性酵母菌吸附铀后—OH的伸缩振动由3434.37 cm-1向低波数移动到3378.65 cm-1,说明—OH参与络合铀酰离子,—OH键键长增长,峰值发生红移,这与刘明学等(2011)的研究结果一致.2100 cm-1左右为C≡C伸缩振动带,吸附铀后峰值向低波数移动了11 cm-1,说明铀酰离子取代了C≡C两端的某些基团,使分子的对称性降低,吸收频率向低频发生偏移(翁诗甫,2010).1643 cm-1处归属于醛羰基CO伸缩振动吸收带,1551 cm-1处的吸收峰为酰胺II带,1242 cm-1为酰胺III带,由N—H键的弯曲振动和C—N键的伸缩振动耦合而成(白静等,2009),在与铀发生相互作用后,这些峰都发生了偏移.综合红外光谱的变化情况来看,在活性酵母菌对铀离子的吸附过程中,羟基、醛羰基、N—H、C—N等为主要的吸附位点.
相对于活性酵母菌,经过高温高压作用的灭活酵母菌在吸附前,2100 cm-1左右的吸收谱带几乎完全消失,蛋白质吸收特征峰(由活性酵母的1643、1551、1242 cm-1分别移动至灭活酵母的1651、1536、1239 cm-1)出现了漂移,说明在高温高压作用下细胞表面结构还是保持完整,只是某些蛋白质类物质变性,C≡C断裂,使谱峰出现了消失和漂移.
从灭活酵母菌吸附前后的红外光谱图(图 7)可看出,在3300~3400 cm4-1之间,峰位由3386.57 cm-1向高波数移动到3434.23 cm-1,可能是由—OH和—NH2的伸缩振动引起的,这与其他学者的发现一致(白静等,2009;Pang et al.,2011).1651 cm-1处归属于酮羰基CO伸缩振动吸收带,1536 cm-1和1239 cm-1是N—H键的弯曲振动和C—N键的伸缩振动,可能有PO伸缩振动的贡献(韩润平等,2000).1072.50 cm-1处是磷酸基—HPO2- 的对称伸缩振动带,与铀离子结合后向低波数偏移了7 cm4-1.因此,灭活酵母菌对铀离子的吸附过程中,羟基、酮羰基、PO、—HPO2-等为主要的吸附位点.
比较图 7中吸附前后活性酵母与灭活酵母的红外光谱可以发现,在918.25 cm-1和916.15 cm-1处为新出现的峰位,这是一个含铀的特征峰,为UO22+的伸缩振动峰(刘明学等,2011;白静等,2009),说明活性酵母与灭活酵母对铀离子确实发生了吸附作用,菌体表面的吸附位点与铀离子形成配合物,表现为表面络合吸附机制.
4 结论(Conclusions)1) 活性酵母与灭活酵母对低浓度含铀溶液都有一定的吸附能力,但灭活酵母菌对低浓度铀的吸附效率明显高于活性酵母菌,且更能适应含低浓度铀的酸性水溶液.
2) 准二级动力学模型的拟合结果表明,活性与灭活酵母在对铀的吸附过程中都存在着电子共用或电子转移过程,这种电荷间的相互作用是一种物理化学现象,说明微生物对铀离子的吸附过程中被动吸附过程为主要的吸附形式.
3) 扫描电镜结果证实了灭活酵母菌的高吸附能力.红外光谱分析进一步说明了活性酵母与灭活酵母菌吸附机理的不同,活性酵母菌对铀离子的吸附过程中,羟基、醛羰基、N—H、C—N等为主要的吸附位点,提供这些吸附位点的主要为多糖和蛋白质类物质;而灭活酵母菌对铀离子的吸附过程中,羟基、酮羰基、PO、—HPO42-等为主要的吸附位点,提供这些位点的主要为多糖和磷酸类物质.说明灭活酵母菌经过高温高压的作用,细胞表面蛋白质类物质变性,细胞膜通透性增加,使其胞内磷酸物质大量释放,大大增加了自身结合铀离子的能力,使灭活酵母菌对铀酰离子的吸附成为一种快速、稳定、高效的吸附过程.而对于活性酵母菌来说,细胞膜的通透性并没有发生改变,在铀离子吸附过程中会选择性吸附,并且细胞自身也会做出不同的调控反应,如在吸附过程中细胞出现凹陷等,这些都说明了活性酵母菌对铀离子的吸附不仅依赖于细胞表面活性位点的被动吸附,同时还要依赖于自身代谢系统的调控及其代谢产物等.
| [${referVo.labelOrder}] | 白静, 秦芝, 王菊芳, 等. 2009. 粘红酵母对铀的吸附研究[J]. 光谱学与光谱分析, 2009, 29(5) : 1218–1221. |
| [${referVo.labelOrder}] | 陈灿, 王建龙. 2006. 酿酒酵母吸附重金属离子的研究进展[J]. 中国生物工程杂志, 2006, 26(1) : 69–76. |
| [${referVo.labelOrder}] | Das N. 2012. Remediation of radionuclide pollutants through biosorption-an overview[J]. Clean-Soil,Air,Water, 40(1) : 16–23. DOI:10.1002/clen.v40.1 |
| [${referVo.labelOrder}] | 代群威, NoonanM J, 董发勤, 等. 2008. 灭活面包酵母菌对溶液中铅离子的吸附研究[J]. 安全与环境学报, 2008, 8(4) : 49–53. |
| [${referVo.labelOrder}] | 代群威, 董发勤, 张伟, 等. 2012. 面包酵母菌对锶离子的吸附行为及其机理研究[J]. 原子能科学技术, 2012, 46(12) : 1423–1428. |
| [${referVo.labelOrder}] | 邓钦文, 王永东, 吕俊文, 等. 2014. 大肠杆菌JM109对废水中铀(VI)的吸附实验研究[J]. 南华大学学报(自然科学版), 2014, 28(1) : 29–33. |
| [${referVo.labelOrder}] | Ding D X, Xin X, Li L, et al. 2014. Removal and recovery of U(VI) from low concentration radioactive wastewater by ethylenediamine-modified biomass of Aspergillus niger[J]. Water Air and Soil Pollution, 225(12) : 1–16. |
| [${referVo.labelOrder}] | Fanhihian H, Peyvandi S. 2012. Adsorption isotherm for uranyl biosorption by Saccharomyces cerevisiae biomass[J]. Journal of Radioanalytical and Nuclear Chemistry, 293(2) : 463–468. DOI:10.1007/s10967-012-1814-y |
| [${referVo.labelOrder}] | Fomina M, Gadd G M. 2014. Biosorption:current perspectives on concept, definition and application[J]. Bioresource Technology, 160 : 3–14. DOI:10.1016/j.biortech.2013.12.102 |
| [${referVo.labelOrder}] | 韩润平, 李建军, 鲍改玲. 2000. 酵母菌吸附铅前后红外光谱比较[J]. 光谱实验室, 2000, 17(4) : 385–387. |
| [${referVo.labelOrder}] | 胡家骏, 周群英. 1998. 环境工程微生物学[M]. . |
| [${referVo.labelOrder}] | 胡恋, 谢水波, 张晓健, 等. 2007. 微生物吸附处理低浓度含铀废水的效能[J]. 安全与环境学报, 2007, 7(2) : 57–60. |
| [${referVo.labelOrder}] | Li L, Ding D X, Hu N, et al. 2014. Adsorption of U(VI) ions from low concentration uranium solution by thermally activated sodium feldspar[J]. Journal of Radioanalytical and Nuclear Chemistry, 299(1) : 681–690. DOI:10.1007/s10967-013-2769-3 |
| [${referVo.labelOrder}] | 林莹, 高柏, 李元峰. 2009. 核工业低浓度含铀废水处理技术进展[J]. 山东化工, 2009, 38(3) : 35–38. |
| [${referVo.labelOrder}] | Liu M X, Dong F Q, Yan X Y, et al. 2010. Biosorption of uranium by Saccharomyces cerevisiae and surface interactions under culture conditions[J]. Bioresource Technology, 101(22) : 8573–8580. DOI:10.1016/j.biortech.2010.06.063 |
| [${referVo.labelOrder}] | 刘明学, 张东, 康厚军, 等. 2011. 铀与酵母菌细胞表面相互作用研究[J]. 高校地质学报, 2011, 17(1) : 53–58. |
| [${referVo.labelOrder}] | Lu X, Zhou X J, Wang T S. 2013. Mechanism of uranium(VI) uptake by Saccharomyces cerevisiae under environmentally relevant conditions:Batch, HRTEM, and FTIR studies[J]. Journal of Hazardous Materials, 262 : 297–303. DOI:10.1016/j.jhazmat.2013.08.051 |
| [${referVo.labelOrder}] | Pang C, Liu Y H, Cao X H, et al. 2011. Biosorption of uranium(VI) from aqueous solution by dead fungal biomass of Penicillium citrinum[J]. Chemical Engineering Journal, 170(1) : 1–6. DOI:10.1016/j.cej.2010.10.068 |
| [${referVo.labelOrder}] | 彭国文, 丁德馨, 胡南, 等. 2012. 纳米Fe3O4负载啤酒酵母菌对铀的吸附性能与机理[J]. 中国有色金属学报, 2012, 22(2) : 605–610. |
| [${referVo.labelOrder}] | Popa K, Cecal A, Drochioiu G, et al. 2003. Saccharomyces cerevisiae as uranium bioaccumulating material:the influence of contact time, pH and anion nature[J]. Nukleonika, 48(3) : 121–125. |
| [${referVo.labelOrder}] | Sarada B, Prasad M K, Kumar K K, et al. 2014. Potential use of Caulerpa fastigiata biomass for removal of lead:kinetics, isotherms, thermodynamic, and characterization studies[J]. Environtal Science and Pollution Research, 21(2) : 1314–1325. DOI:10.1007/s11356-013-2008-z |
| [${referVo.labelOrder}] | Vijayaraghavan K, Yun Y S. 2008. Bacterial biosorbents and biosorption[J]. Biotechnology Advances, 26(3) : 266–291. DOI:10.1016/j.biotechadv.2008.02.002 |
| [${referVo.labelOrder}] | Wang J L, Chen C. 2006. Biosorption of heavy metals by Saccharomyces cerevisiae:A review[J]. Biotechnology Advances, 24(5) : 427–451. DOI:10.1016/j.biotechadv.2006.03.001 |
| [${referVo.labelOrder}] | Wang J L, Chen C. 2009. Biosorbents for heavy metals removal and their future[J]. Biotechnology Advances, 27(2) : 195–226. DOI:10.1016/j.biotechadv.2008.11.002 |
| [${referVo.labelOrder}] | 魏广芝, 徐乐昌. 2007. 低浓度含铀废水的处理技术及其研究进展[J]. 铀矿冶, 2007, 26(2) : 90–95. |
| [${referVo.labelOrder}] | 翁诗甫. 2010. 傅里叶变换红外光谱分析(第2版)[M]. 北京: 化学工业出版社 : 300–301. |
| [${referVo.labelOrder}] | 吴海云, 米娴妙, 冯仕训, 等. 2010. 啤酒酵母菌吸附Cr(VI)的动力学及热力学研究[J]. 应用化工, 2010, 39(4) : 532–537. |
| [${referVo.labelOrder}] | 夏良树, 谭凯旋, 邓帛辉, 等. 2008. 四种生物吸附剂对于铀的吸附性能研究[J]. 化工工程, 2008, 36(2) : 9–12. |
| [${referVo.labelOrder}] | Xiao X L, Xue J H, Ding D X, et al. 2016. Adsorption of low concentration of uranium(VI) from aqueous solution by diethylenetriamine functionalized cycas revolute leaves[J]. Journal of Radioanalytical Nuclear Chemistry, 308(3) : 1027–1037. DOI:10.1007/s10967-015-4601-8 |