 2017, Vol. 37
2017, Vol. 37



2. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306;
3. 江苏省海洋水产研究所, 南通 226007
2. Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306;
3. Maine Fisheries Research Institution of Jiangsu, Nantong 226007
浮游植物是海洋环境中重要的生物组成类群之一,是水体中的初级生产力也是初级消费者的摄食来源(左涛等,2008),在食物网能量流动等生态过程中起着重要作用(张婷等,2014).浮游植物种类及其物种多样性不仅可用来描述群落结构,也可以反映水体的营养状况(夏品华等,2011),但其与环境因子在时间和空间尺度上关系复杂,还没有一致的理论解释(Reynolds,1998).目前用于浮游植物群落与环境因子动态分析的统计方法主要有主成分分析(PCA)、典范对应分析(CCA)等(Abrantes et al.,2006;Bernard et al.,2004;Daniela et al.,2006;李秋华等,2007),其中,典范对应分析可以将多个相关的环境因子结合,分析其间的复杂关系,并能准确直观地反映出来,是研究浮游植物群落和环境因子关系的有效工具(王宇飞等,2015).近些年,国内学者对典范对应分析的研究集中在淡水湖泊与水库区域(沈会涛等,2008;夏品华等,2011),而近海岛屿及海洋牧场等以海洋为主的区域的研究相关较少.
海州湾海洋牧场的建设促进了当地养殖业的发展,改善了该海域的渔业资源和生态环境状况(张虎等,2005).本文以连云港海州湾海洋牧场海域2013年环境因子和浮游生物采样调查为基础,采用群落多样性指数统计、聚类分析和CCA等方法研究海洋牧场区浮游植物群落结构特征,探讨海洋牧场区浮游植物季节变动规律及与环境因子之间的关系,以期为海州湾海洋牧场海域的生态环境保护和评价提供参考.
2 材料与方法(Materials and methods) 2.1 研究区域分别于2013年春(5月)、夏(8月)、秋(10月)3个季节对海州湾海洋牧场海域进行了环境因子和浮游生物的调查,每次调查选择23个站点,其中,人工鱼礁区17个(R1~R17),对照区6个(C1~C6),对照区设置在远离人工鱼礁区4海里外的海域,调查范围为34°52.15′~34°58.00′N,119°25.75′~119°30.517′E,采样站点如图 1所示.
 |
| 图 1 海州湾采样站点图 Fig. 1 locations of Sampling stations in Haizhou Bay |
浮游植物样品的采集采用浅水Ⅲ型浮游生物网(网口直径为37 cm,网目孔径0.077 mm),用绞车自海底到水面垂直拖取,起网速度为0.5 m·s4-1.采集的样品用缓冲甲醛溶液固定保存,固定好的样品带回实验室进行种类鉴定和数量统计.水环境监测内容同步开展,包括底泥中的总氮(TN)、总磷(TP)及溶解氧(DO)、生化需氧量(BOD5)、化学需氧量(CODMn)、氨氮(NH4+-N)、亚硝酸盐氮(NO2--N)、硝酸盐氮(NO3--N)、硅酸盐(SiO32--Si)、活性磷酸盐(PO43--P)、叶绿素a(Chl-a)、悬浮物(SS)、水温(SST)、透明度(TRA)等,底泥使用抓斗式采泥器采集,各类营养盐通过采集各站点1 L海水水样,装入至聚乙烯瓶,并加入分析纯级氯仿进行固定;叶绿素a采用分光光度法测定,通过各站点采集500 mL海水,并加入5 mL碳酸镁,经过 0.45 μm孔径的微孔滤膜抽滤获得;悬浮物通过各站点采集500 mL海水,经过 0.45 μm孔径的预称重的微孔滤膜抽滤获得;水温用表层水温温度计测定,透明度用透明度盘测定,样品均冷冻保存带回实验室处理.样品的采集、保存及分析按中华人民共和国国家标准《海洋调查规范》(GB12763—2007)进行.
2.3 数据处理在实验室进行种类鉴定和分析,浮游植物的定性采用显微镜(OLYMPUS BX35)镜检方式,定量检测方法参照GB12713.6—2007,采用0.1 mL的浮游植物记数框进行种类的鉴定和丰度的分析.以浮游植物种类鉴定、计数及分度的统计数据为基础,对每次调查的人工鱼礁区、对照区的浮游植物生物多样性进行进一步分析,主要选用辛普森(Simpson)多样性指数(D)、Shannon-wiener指数(H′)、均匀度指数(J)、优势度Y等参数(沈国英等,2002);计算公式如下:
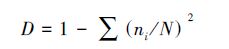
|
(1) |
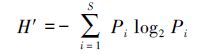
|
(2) |
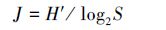
|
(3) |

|
(3) |
式中,ni为第i种的个体数,N为出现的总个体数,S为样品中的种类总数,Pi为第i种的个体数(ni)与总个体数(N)的比值(ni/N)
根据优势度指数的计算,选定优势度指数大于0.02的种类为优势种;数据处理及绘图采用Excel2007、SPSS13、Primer5.0及Gri等统计分析软件.另外,采用Canoco for Windows5.0软件对浮游植物物种和环境因子进行典范对应分析.
3 结果与分析(Results and analysis) 3.1 环境因子春、夏和秋季海州湾海洋牧场各环境因子数据如表 1所示,SiO32--Si、NO3--N、NO2--N、BOD5和SS等5个环境因子平均值春季最大,其中,SiO32--Si、NO3--N、BOD5平均值对照区要高于人工鱼礁区;Chl-a、COD、TP和TN平均值夏季均高于春、秋两季,其中,Chl-a和TP浓度对照区高于人工鱼礁区.
| 表 1 海州湾海洋牧场环境因子特征 Table 1 Distribution of environmental factors in Haizhou Bay |
海州湾浮游植物种类较多,2013年3次跟踪调查中,鉴定到浮游植物种类共115种,其中,硅藻门(Bacillariophyta)97 种,甲藻门(Pyrrophyta)16种,蓝藻门(Cyanophyta)1种,绿藻门(Chlorophyta)1种.从海州湾海洋牧场各季节浮游植物种类数和丰度的平面分布图(图 2)可以看出,春季浮游植物丰度在中部海域较高,主要集中在人工鱼礁区,夏季丰度在南部海域较高,秋季丰度较大值也集中在人工鱼礁区,尤其在东南部海域,峰值达到19.41×104 cells·m-3,随着季节变化,较高丰度区域有向低纬度迁移的趋势;各个季节的浮游植物种类数在东部海域较高,最大值为秋季的36种.
 |
| 图 2 海州湾海洋牧场各季节浮游植物丰度及种类数分布图 Fig. 2 distribution of phytoplankton abundance and taxa in different seasons of Haizhou Bay |
人工鱼礁区和对照区在不同季节的浮游植物种类数上相差较小,无明显差异(p>0.05)(表 2).人工鱼礁区和对照区浮游植物种类数均值的季节变化均表现为秋季(约30种)高于夏季(约10种)高于春季(约5种)的趋势.浮游植物丰度最大值为秋季RA14站点的19.68×104 cells·m-3,最小值为春季RA11站点的3.29×104 cells·m-3;人工鱼礁区浮游植物平均丰度大小为秋季>夏季>春季,对照区为秋季>春季>夏季,人工鱼礁区3个季节的浮游植物丰度都明显高于对照区(p<0.05)(表 3),人工鱼礁区和对照区的浮游植物丰度和种类数均值均为秋季最高.
| 表 2 不同区域浮游植物群落种类数的多重方差检验 Table 2 Multiple comparison test for taxa of phytoplankton community in different areas |
| 表 3 不同区域浮游植物群落丰度的多重方差检验 Table 3 Multiple comparison test for abundance of phytoplankton community in different areas |
海州湾人工鱼礁区5月优势种主要是星脐圆筛藻(Coscinodiscus asteromphalus)、佛氏海线藻(Thalassiothrix frauenfeldii)、虹彩圆筛藻(Coscinodiscus oculusiridis)等,8月主要是爱氏辐环藻(Coscinodiscus oculusiridis)、菱形海线藻(Thalassionema nitzschioides)、夜光藻(Noctiluca scintillans)等,10月主要是条纹小环藻(Cyclotella striata)、叉状角藻(Ceratium furca)、环纹娄氏藻(Lauderia annulatus);对照区5月主要是星脐圆筛藻(Coscinodiscus asteromphalus)、爱氏幅环藻(Coscinodiscus oculusiridis)、细弱圆筛藻(Coscinodiscus subtilis)等,8月主要是佛氏海线藻(Thalassiothrix frauenfeldii)、叉状角藻(Ceratium furca)、夜光藻(Noctiluca scintillans)等,10月主要是宽笔尖形根管藻(Rhizosolenia styliformis)、环纹娄氏藻(Lauderia annulatus)、条纹小环藻(Cyclotella striata)等.
从表 4中可以看出,海州湾人工鱼礁区浮游植物群落辛普森多样性指数D平均值的范围是0.640~0.892,最高值0.892出现在10月,最低值0.640出现在8月,对照区的最低值0.662也出现在8月,最高值0.836同样出现在10月,其范围为0.662~0.836;香农-威纳指数平均值在人工鱼礁区范围是1.996~3.312,最大值出现在10月,最小值出现在8月,对照区平均值范围是2.056~3.109,最小值出现在5月,最大值出现在10月;均匀度J在人工鱼礁区的平均值范围是0.628~0.908,其最大值出现在5月,最小值出现在8月,对照区的平均值范围是0.709~0.910,最大值出现在5月,最小值出现在8月,可以看出,多样性指数在人工鱼礁区和对照区存在时空差异.
| 表 4 海州湾浮游植物群落多样性指数 Table 4 Diversity index of phytoplankton community |
根据Primer5.0软件分析不同季节及区域的浮游植物群落(图 3),从分析结果中可以看出,春季浮游植物群落组成和夏、秋两季存在不同,春季人工鱼礁区和对照区的相似度较高,夏季人工鱼礁区和秋季人工鱼礁区的浮游植物群落组成相似,夏季对照区和秋季对照区相似.从对不同季节的浮游植物群落丰度多重比较方差分析中看出(表 5、表 6),秋季浮游植物丰度和种类数均明显大于夏季和春季(p<0.05),而春季小于秋季和夏季(p<0.05).
 |
| 图 3 不同季节及区域浮游植物群落的聚类分析 Fig. 3 Cluster analysis based on Bray-Curtis similarity matrix of phytoplankton community in different seasons and areas |
| 表 5 不同季节浮游植物群落丰度的多重方差检验 Table 5 Multiple comparison test for abundance of phytoplankton community in different seasons |
| 表 6 不同季节浮游植物群落种类数的多重方差检验 Table 6 Multiple comparison test for taxa of phytoplankton community in different seasons |
运用Canoco for Windows5.0软件对浮游植物数据和环境数据进行典范对应分析.经条件筛选,用于排序的环境因子如表 7所示,在CCA排序图中TN与第一排序轴呈最大正相关,且相关系数为0.8074,其次为SiO32--Si,相关系数为0.7930,COD与第一排序轴呈最大负相关,相关系数为-0.6626,其次为PO43--P,相关系数为-0.6593;与第二排序轴呈最大正相关的环境因子为生物需氧量,相关系数为0.7548,其次为NH4+-N,相关系数为0.4195,而与第二排序轴呈最大负相关的环境因子为TP,相关系数为-0.4770.用于排序的浮游植物种名录见表 8.
| 表 7 环境因子与两个排序轴间的相关系数 Table 7 Correlation coefficients of environmental factors and two axes of CCA |
| 表 8 用于CCA排序的浮游植物名录 Table 8 The list of phytoplankton for CCA |
从图 4的浮游植物和环境因子的二维排序图中可以看出,左上方这些物种之间的距离较小,这说明它们之间的分布差异程度小,SiO32--Si、NO3--N、TN对其影响较大.其中,日本角毛藻(Chaetoceros nipponica)及肾形圆筛藻(Coscinodiscus reniformis)呈显著正相关(p<0.05),主要分布于NO3--N含量高的海水中,同时这几个物种对于TN的需求量也较大.位于右上方的有根管藻sp.(Rhizosolenia sp.)、细弱圆筛藻(Coscinodiscus subtilis)、中华根管藻(Rhizosolenia sinensis)、海链藻sp.( Thalassiosira sp.)、圆海链藻(Thalassiosira rotula),这些物种之间的距离相对较大,这说明它们之间的分布差异程度较大,其中,海链藻sp.( Thalassiosira sp.)、圆海链藻(Thalassiosira rotula)对COD的需求要高于剩下的3种藻类.位于左下方的双脊角毛藻(Chaetoceros costatus)、丹麦细柱藻(Leptocylindrus danicus)、弓束圆筛藻(Coscinodiscus curvatulus)、虹彩圆筛藻(Coscinodiscus oculusiridis)、尖刺伪菱形藻(Pseudonitzschia pungens)、柔弱伪菱形藻(Leptocylindrus danicus)、格氏圆筛藻(Coscinodiscus hainanensis)对NH4+-N的需求较高,同时它们的分布与BOD密切相关.位于右下方的有宽笔尖形根管藻(Rhizosolenia styliformis var)、小环藻sp.(Cyclotella sp.)、条纹小环藻(Cyclotella striata)、针杆藻sp.(Synedra sp.)、太阳双尾藻(Ditylum sol)、双角马鞍藻(Campylodiscus biangulatus)、曲舟藻sp.(Pleurosigma sp.),对总TP的需求较高,且相互的距离较近,说明它们之间的分布差异较小.
 |
| 图 4 海州湾海洋牧场浮游植物物种与环境因子关系的二维排序图 Fig. 4 CCA biplot of phytoplankton species and environmental factors in marine ranching of Haizhou Bay |
2013年春、夏、秋3季共记录浮游植物115种,其中,硅藻门占84.3%,硅藻门种类较多与该海域水体中存在的营养盐有着密切联系,2009年调查海州湾海域时就发现,硅酸盐在水体中含量较高,并且它是促进硅藻生长的主要营养盐之一(杨柳等,2011),这与本次调查相一致.海洋牧场秋季浮游植物丰度及种类数均要远高于春、夏两季,这与杨晓改等(2014)在2011年在海州湾及其附近海域调查时得到的结论一致.春、秋两季是赤潮爆发的高峰期,浮游植物丰度高于夏季(张永山等,2002),夏季水温较高也会抑制浮游植物生长(江志兵等,2012).但此次调查结果显示,春季浮游植物丰度及种类数少于夏季,出现这种情况可能由于当年夏季降雨量较大,受地表径流和入海河流携带影响,使水体中的营养盐含量增加,导致浮游植物丰度增加(高春梅等,2015).
从浮游植物群落多样性指数的分析结果来看,辛普森多样性指数D主要判断群落物种多样性,香农-威纳指数可以反映浮游植物群落的多样性和复杂程度,辛普森多样性指数和香浓-威纳指数结合使用,均匀度反映了浮游植物的均匀分配情况,许晴等(2011)通过假设的群落条件及情景设计证明,前者辛普森多样性指数比香农-威纳指数对均匀度更为敏感,而香浓-威尔指数对物种丰富度更敏感.浮游植物的辛普森多样性指数及香农-威纳指数均以秋季最高,秋季物种较春、夏两季更为丰富而且群落结构更为复杂,说明海区内秋季的环境状况更加适宜于浮游植物的生长和繁殖.杨晓改等(2014)在2011年对海州湾及其附近海域调查时发现,香农-威纳指数、均匀度指数和辛普森多样性指数D 均为夏、秋季高,冬、春季低,这与本文研究结果一致,在秋季多样性指数均高于春、夏两季.聚类分析结果是对物种参数进行log(x+1)转换和标准化后,建立Bray-Curtis相似性聚类分析,是多种参数耦合的结果.聚类分析结果显示,浮游植物群落组成在春季和夏、秋两季有明显的区别,并且春季人工鱼礁区和对照区的浮游植物群落组成上无明显变化,说明春季人工鱼礁区和对照区浮游植物群落组成相似,而夏、秋两季浮游植物群落组成存在差异.江志兵等(2012)在象山港人工鱼礁区对浮游植物群落聚类分析时发现,各个季节鱼礁区与对照区在浮游植物群落组成上相似;但夏、秋两季中人工鱼礁区和对照区的浮游植物群落组成上在区域上无差异.一方面在浮游植物群落参数的选择,本研究是利用多样性指数进行分析耦合,而江志兵等(2012)是利用丰度等参数进行分析;在时间尺度上,本研究春季人工鱼礁区和对照区浮游植物群落组成相似,而夏、秋两季浮游植物群落组成存在差异,而江志兵等(2012)发现各个季节的人工鱼礁区及对照区都发现浮游植物组成相似度较高.
从浮游植物与两个排序轴的相关系数来看,总氮、硅酸盐和总磷是影响浮游植物分布的主要环境因子,本文中总氮和总磷来自于表层沉积中,其对浮游植物的影响主要是通过向上覆水释放营养盐而产生作用.韩照祥等(2012)的研究也表明,底泥中的营养盐会在生物、化学和物理等一系列作用下,向上覆水释放营养盐,从而使沉积物中营养盐从固相转化成液相,溶解在水体中,间接影响到浮游植物.近年来,高春梅等(2015)利用实验室培养法,研究了海州湾海洋牧场沉积物-水界面的交换状况,结果表明,无机氮在春、夏、秋3季均由沉积物向水体进行迁移,而且营养盐在夏季和秋季向水体释放;而本研究表明,海州湾游植物种类数和丰度的季节变化表现为秋季>夏季>春季的趋势,因此,夏季和秋季浮游植物生长对水体中营养盐消耗较大,因而需要从沉积物向水体中进行营养盐补充,这也进一步印证了上述研究结果.硅酸盐是也是影响浮游植物分布的主要环境因子之一,Si是硅酸盐中重要的元素,也是硅藻细胞壁的主要成分,所以硅酸盐是硅藻必需的营养盐类(张硕等,2006);Si不会对整个浮游植物群落产生限制作用,但会影响浮游植物群落的组成(王勇等,2000).海水中的硅主要来源于河水和其它陆地径流,沿岸海域由于河水的补给一般不缺少 Si,并使 Si∶N或Si∶P保持一定比值,一般Si∶N较高时硅藻占优势,Si∶N较低时甲藻或其他鞭毛藻占优势,硅藻需要的Si∶N 原子比约为1∶1(Redfieid et al.,1963).硅酸盐对硅藻的生长有比较大的影响,在海州湾浮游植物群落中,硅藻占84.3%,间接地对整个浮游植物群落组成造成影响.
此外,浮游植物群落结构与环境因子的关系也与所处区域密切相关,景明等(2014)对阳澄湖浮游植物调查时发现,总磷、氨氮和水温是影响浮游植物群落结构的主要环境因子;栾青杉等(2007)发现,透明度、硝酸盐和硅酸盐是影响长江口海域的浮游生物群落结构的主要环境因子.在海州湾海域,杨晓改等(2014)于2011年进行大面积综合调查时发现,水表面温度(SST)、营养盐和溶解氧(DO)是影响浮游植物丰度和分布的主要环境因子,研究结果与本研究有所差别,这可能与调查区域有关,本研究主要集中在海州湾海洋牧场区域,人工鱼礁比较集中并且会改变局部生境,导致环境变化.江志兵等(2012)在2010年对象山港人工鱼礁区及其邻近海域浮游植物调查时发现,区域间(人工鱼礁区和对照区)基本无显著差异,这与本研究存在差异,海州湾海洋牧场区从2003年开始投放人工鱼礁,至今已经达到一定规模,并且人工鱼礁局部生境相对稳定,而象山港人工鱼礁投放数量不多,建礁时间短,导致其生态效应在短期内难以显著体现;其次本研究对照区采样站点选取离人工鱼礁区4海里外的天然海域,而象山港调查选点距离较近,且采样点均靠近岛屿,潮流和岛屿对浮游植物的影响可能超过了人工鱼礁投放对其影响.
5 结论(Conclusions)1) 2013年春、夏、秋3季海州湾海洋牧场浮游植物群落调查中,硅藻门是主要优势种,秋季的浮游植物种类数和丰度均高于春、夏两季.
2) 秋季浮游植物群落的辛普森多样性指数和香浓-威尔指数较高,人工鱼礁区春、秋两季浮游植物群落结构与对照区相比更为复杂.
3) 方差分析和聚类分析结果,海州湾海洋牧场浮游植物群落存在时空差异,春季浮游植物群落丰度明显高于夏、秋两季(p<0.05),人工鱼礁区和对照区也存在明显季节性差异(p<0.05).
4) 总氮、硅酸盐和总磷是影响海州湾海洋牧场浮游植物种群分布的主要环境因子,对海州湾浮游植物优势种影响最大的是硅酸盐.
| [${referVo.labelOrder}] | Abrantes N, Antunes S C, Pereira M J, et al. 2006. Seasonal succession of cladocerans and phytoplanktonand their interactions in a shallow eutrophic lake(Lake Vela,Portugal)[J]. Acta Oecologica, 29(1) : 54–64. DOI:10.1016/j.actao.2005.07.006 |
| [${referVo.labelOrder}] | 高春梅, 朱珠, 王功芹, 等. 2015. 海州湾海洋牧场海域表层沉积物磷的形态与环境意义[J]. 中国环境科学, 2015, 35(11) : 3437–3444. |
| [${referVo.labelOrder}] | 高春梅, 郑伊汝, 张硕. 2016. 海州湾海洋牧场沉积物-水界面营养盐交换通量的研究[J]. 大连海洋大学学报, 2016, 31(1) : 95–102. |
| [${referVo.labelOrder}] | 国家质量监督检验检疫总局.2007.GB/T12763.4-2007海洋调查规范第6部分:海洋生物调查[S].北京:中国标准出版社 |
| [${referVo.labelOrder}] | 韩照祥, 王超, 朱圳, 等. 2012. 海州湾及其临近海域沉积物-水界面的磷形态特征与动力学[J]. 环境科学与技术, 2012, 35(11) : 12–15. |
| [${referVo.labelOrder}] | 景明, 李继影, 刘孟宇, 等. 2014. 阳澄湖浮游植物群落结构与环境因子的典范对应研究[J]. 环境科学与管理, 2014, 39(11) : 145–148. |
| [${referVo.labelOrder}] | 姜勇, 许恒龙, 朱明壮, 等. 2010. 胶州湾浮游原生生物时空分布特征-丰度周年变化及与环境因子间的关系[J]. 中国海洋大学学报(自然科学版), 2010, 40(3) : 17–23. |
| [${referVo.labelOrder}] | 江志兵, 陈全震, 寿鹿, 等. 2012. 象山港人工鱼礁区的网采浮游植物群落组成及其与环境因子的关系[J]. 生态学报, 2012, 32(18) : 5813–5824. |
| [${referVo.labelOrder}] | 孔苗苗.2013.岩滩水库浮游生物群落结构及水质评价[D].南宁:广西大学.1-746 |
| [${referVo.labelOrder}] | 林青, 由文辉, 徐凤洁, 等. 2014. 滴水湖浮游动物群落结构及其与环境因子的关系[J]. 生态学报, 2014, 34(23) : 6918–6929. |
| [${referVo.labelOrder}] | 李秋华, 韩博平. 2007. 基于CCA的典型调水水库浮游植物群落动态特征分析[J]. 生态学报, 2007, 27(6) : 2355–2364. |
| [${referVo.labelOrder}] | 栾青杉, 孙军, 宋书群, 等. 2007. 长江口夏季浮游植物群落与环境因子的典范对应分析[J]. 植物生态学报, 2007, 31(3) : 445–450. |
| [${referVo.labelOrder}] | Parinet B, Lhote A, Legube B. 2004. Principal component analysis:an appropriate tool for water quality evaluation and management-application to a tropical lake system[J]. Ecological Modelling, 178(1) : 295–311. |
| [${referVo.labelOrder}] | Redfieid A C, Ketchum B H, Richards F A. 1963. The Influence of Organism on the Composition of Sea Water[M]. New York: Interscience Publishers . |
| [${referVo.labelOrder}] | Reynolds. 1998. What factors influence the species composition of phytoplanktonin lakes of different trophic status?[J]. Hydrobiologia, 370(1) : 11–26. |
| [${referVo.labelOrder}] | 沈国英, 施并章. 2002. 海洋生态学[M]. 北京: 科学出版社 : 1–446. |
| [${referVo.labelOrder}] | 沈会涛, 刘存歧. 2008. 白洋淀浮游植物群落及其与环境因子的典范对应分析[J]. 湖泊科学, 2008, 20(1) : 773–779. |
| [${referVo.labelOrder}] | 王勇, 崔念志. 2000. 营养盐对浮游植物生长上行效应机制的研究进展[J]. 海洋科学, 2000, 24(10) : 30–32. |
| [${referVo.labelOrder}] | 王宇飞, 赵秀兰, 何丙辉, 等. 2015. 汉丰湖夏季浮游植物群落与环境因子的典范对应分析[J]. 环境科学, 2015, 36(3) : 922–927. |
| [${referVo.labelOrder}] | 夏品华, 马健荣, 李存雄, 等. 2011. 红枫湖水库冬春季浮游生物群落与环境因子的典范对应分析[J]. 环境科学研究, 2011, 24(4) : 18–26. |
| [${referVo.labelOrder}] | 许晴, 张放, 许中旗, 等. 2011. Simpson指数和Shannon-Wiener指数若干特征的分析及"稀释效应"[J]. 草业科学, 2011, 28(4) : 527–531. |
| [${referVo.labelOrder}] | 杨柳, 张硕, 孙满昌, 等. 2011. 海州湾人工鱼礁海域春、夏季浮游植物群落结构及其与环境因子的关系[J]. 生物学杂志, 2011, 28(6) : 14–18. |
| [${referVo.labelOrder}] | 杨晓改, 薛莹, 昝肖肖, 等. 2014. 海州湾及其邻近海域浮游植物群落结构及其与环境因子的关系[J]. 应用生态学报, 2014, 25(7) : 2123–2131. |
| [${referVo.labelOrder}] | 张虎, 朱孔文, 汤建华. 2005. 海州湾人工鱼礁养护资源效果初探[J]. 海洋渔业, 2005, 27(1) : 38–43. |
| [${referVo.labelOrder}] | 张硕, 朱孔文, 孙满昌. 2006. 海州湾人工鱼礁区浮游植物的种类组成和生物量[J]. 大连水产学院学报, 2006, 21(2) : 134–140. |
| [${referVo.labelOrder}] | 张婷, 马行厚, 王桂苹, 等. 2014. 鄱阳湖国家级自然保护区浮游生物群落结构及空间分布[J]. 水生生物学报, 2014, 38(1) : 160–167. |
| [${referVo.labelOrder}] | 张永山, 吴玉霖, 邹景忠, 等. 2002. 胶州湾浮动弯角藻赤潮生消过程[J]. 海洋与湖沼, 2002, 33(1) : 55–61. |
| [${referVo.labelOrder}] | 左涛, 王俊, 金显仕, 等. 2008. 春季长江口邻近外海网采浮游生物的生物量谱[J]. 生态学报, 2008, 28(3) : 293–301. |