 2017, Vol. 35
2017, Vol. 35扩展功能
文章信息
- 陈应权, 马利涛, 彭琳, 刘军, 张立, 范俊杰, 闫德飞
- CHEN YingQuan, MA LiTao, PENG Lin, LIU Jun, ZHANG Li, FAN JunJie, YAN DeFei
- 甘肃宝积山盆地中侏罗世阿干镇似银杏(Ginkgoites aganzhenensis Yang)的古环境意义
- The Paleoenvironmental Significance of Ginkgoites aganzhenensis Yang from the Middle Jurassic in the Baojishan Basin, Gansu Province
- 沉积学报, 2017, 35(1): 57-66
- ACTA SEDIMENTOLOGICA SINCA, 2017, 35(1): 57-66
- 10.14027/j.cnki.cjxb.2017.01.006
-
文章历史
- 收稿日期:2016-01-08
- 收修改稿日期: 2016-05-20




2. 甘肃省西部矿产资源重点实验室, 兰州 730000;
3. 武警黄金地质研究所, 河北廊坊 065000
2. Key Laboratory of Mineral Resources in Western China (Gansu Province), Lanzhou University, Lanzhou 730000, China;
3. Gold Geological Institute of CAPF, Langfang, Hebei 065000, China
20世纪以来,温室效应所带来的全球变暖已成为全人类最关注的环境问题之一,而CO2作为一种主要的温室气体,影响着全球气候的变化[1]。因此,探索地史时期的CO2浓度与气候的关系,对于认识今天大气CO2浓度的变化、更有效的预测气候的未来发展方式具有重要的意义。
植物是指示气候与生存环境之间的良好指标[2-3],气孔作为陆生植物与大气交换水分和CO2的通道,它的分布、数目和密度等特征与当时的大气CO2等环境信息密切相关,通过对载有气孔的植物角质层的各项特征研究可推测当时外界环境的变化。探索不同地史时期植物与环境之间的内在联系,利用植物化石获取古大气CO2浓度的变化信息,已成为当今古植物学研究的一个重要内容[4-5]。自Woodward[3]于1987年首次发现了植物叶片气孔参数与CO2浓度的负相关关系后,科学家们基于这种关系对新生代、中生代,甚至古生代的古大气CO2浓度变化情况进行了分析[6-10]。银杏类(Ginkgoalean)植物由于其形态学上的“迟滞性”[11-12],已成为科学家们定量重建古气候、古环境的重要手段,选用此类化石进行不同地史时期古大气CO2浓度的重建也得到了广泛的应用[13-22]。
本文报道了产于甘肃宝积山盆地中侏罗统的阿干镇似银杏(Ginkgoites aganzhenensis),研究了其角质层微细构造,统计其气孔参数,并使用气孔指数法和气孔比率法对研究区中侏罗世早期的大气CO2浓度进行了重建,对两种方法进行比较。将其与其他同期的相关资料进行对比,分析不同种类植物化石在重建古大气CO2浓度时的差异性, 进而通过其气孔参数重建古温度,分析化石生长时期的古环境。
1 区域地理地质概况宝积山盆地位于甘肃省白银市平川区,其分布范围大致为103°20'~105°00' E,36°00'~36°50' N。由于晚三叠世印支运动引起的海退以及北半球潮湿气候带的北移,我国早、中侏罗世聚煤盆地以内陆湖盆为主并主要发育于西北和华北地区[23]。宝积山盆地就是形成于该时期的具代表性的坳陷型聚煤盆地之一,其位于祁连山东端,在地史时期受秦祁褶皱强烈影响,盆地内断层等构造较发育[24-25]。盆地内从寒武系到第四系都有出露,以显生宇发育较为完整[24, 26]。盆地内中生界发育,产出较丰富的植物化石,化石采集位置如图 1。

|
| 图 1 甘肃宝积山盆地化石采集点位置图 Figure 1 Geographical location of the Baojishan Basin, Gansu province |
化石产出层位为中侏罗统窑街组(J2y),其主要出露于宝积山向斜两翼,为区内主要含煤地层。该组上覆中侏罗统新河组(J2x),下伏下侏罗统大西沟组(J1d),且均为平行不整合关系(图 2)。岩性主要以深灰色页岩,灰色、灰白色砂岩,灰白色石英砂岩、石英砾岩为主,局部夹煤层。Ginkgoites aganzhenensis采于该组含煤岩段的灰黑色炭质泥岩中。窑街组在宝积山盆地的总厚度为77~279 m (图 2)。该组含较丰富的植物化石,其组合为典型的奇丽锥叶蕨-似银杏组合(Coniopteris spectabilis-Ginkgoites),与兰州窑街煤田中侏罗世早期的膜蕨型锥叶蕨-似银杏组合(Coniopteris hymenophylloides-Ginkgoites)接近,其地质年代为中侏罗世早期,相当于阿林期(Aalenian)-巴柔期(Bajocian)[27-28]。

|
| 图 2 宝积山盆地窑街组柱状图 Figure 2 The stratigraphic column of the Yaojie Formation in the Baojishan Basin |
化石角质层处理方法:首先选取角质层保存较完整的植物化石标本进行拍照,局部处理后,取角质层样。样品经蒸馏水浸润并洗涤后,先后用10%的HCl溶液与50%的HF溶液处理以除去碳酸盐类物质以及硅酸盐类物质,后用蒸馏水冲洗至中性并将其放于舒氏液中,角质层颜色变为淡黄色或近透明状时用蒸馏水冲洗至中性。再经氨水,后用蒸馏水处理至中性,在体视显微镜下观察并分离出上、下表皮,将一部分处理好的角质层用番红溶液进行染色后制成光学薄片,在Leica DM4000显微镜下观察和照相。另一部分样品用酒精溶液梯度脱水,用导电双面胶粘在铜座上喷金,使用JEOL-JSM-5600LV扫描电子显微镜对其进行观察、照相。
2.2 两种基于气孔的古大气CO2浓度重建方法气孔指数SI(stomatal index)是指1 mm2叶片上气孔数除以气孔数与普通表皮细胞数之和,用百分比表示(%),计算公式如下:
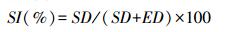 (1)
(1)
其中,SD为气孔数,ED为普通表皮细胞数。
2.2.1 气孔指数法这是一种定量的方法,通过“温室实验”结合腊叶标本与冰芯记录,归纳出现生种气孔指数与大气CO2浓度之间的函数关系(Regression Function, RF),将该函数应用于相应的化石种对当时的古大气CO2浓度进行重建[7, 16]。特别是Royer et al.[7]归纳出了现生银杏气孔参数与大气CO2环境之间的回归函数关系,并利用化石材料对新生代的古大气CO2变化进行了重建。本文选用Royer et al.[7]所建立的经验公式:
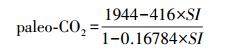 (2)
(2)
其中paleo-CO2为古大气CO2浓度,单位为×10-6。
2.2.2 气孔比率法气孔比率SR(stomatal ratio)是指化石植物的“最近现生对应种”(the Nearest living equivalent species, NLE)或“最近现生亲缘种”(the Nearest living relative species, NLR)的气孔指数与化石气孔指数的比值[29]。然后将气孔比率按照一定的标准转化为全球碳平衡模型中的RCO2,计算古大气CO2浓度值。利用气孔比率法恢复古大气CO2浓度有两种标准[8, 30-31]:一种是石炭纪标准,主要用于古近纪以前不同地史时期古大气CO2浓度的重建,1个SR单位相当于2个RCO2单位;一种是最近评估标准,主要用于新生代以来的古大气CO2浓度的重建,1个SR单位相当于1个RCO2单位。RCO2与古大气CO2浓度之间的转换关系为:
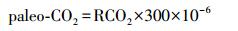 (3)
(3)
当前化石采于中侏罗统,故采用石炭纪标准。
3 植物化石鉴定特征银杏目Ginkgoales
银杏科Ginkgoaceae
似银杏属Ginkgoites Seward 1919, emend. Florin 1936
阿干镇似银杏Ginkgoites aganzhenensis Yang 1988
1988.Ginkgoites aganzhenensis,Yang,p.73, pl.1, figs. 1~4。
宏观特征:标本共三块,为叶片压型化石。叶大,具柄,柄长至少2.5 cm,宽约2 mm。叶片高约10.8 cm,宽14 cm,先深裂为左右两半,每一半再深裂1~3次,形成4~8个裂片。裂片为梭形,顶端尖圆,基部急剧狭缩,彼此分离。裂片最长可达10.8 cm,最宽处1.8 cm。裂片长度有从中间向两侧减短的趋势。叶脉每厘米约10条,在基部作二歧式分叉,中上部不分叉,裂片中部平行,于顶部略聚敛(图 3)。

|
| 图 3 宝积山盆地Ginkgoites aganzhenensis植物化石 a. Ginkgoites aganzhenensis标本(标本号:GSPC-LDGSW-2014-001);b.图a的素描图;c. Ginkgoites aganzhenensis标本(标本号:GSPC-LDGSW-2014-002),示叶柄;d. Ginkgoites aganzhenensis标本(标本号:GSPC-LDGSW-2014-003);e.示叶脉特征。 Figure 3 Fossil specimens of Ginkgoites aganzhenensis from the Baojishan Basin |

|
| 图 4 Ginkgoites aganzhenensis表皮特征 a.下表皮内表面(SEM),箭头所指为气孔;b.下表皮外表面(SEM),箭头所指为气孔;c.下表皮内表面(LM),箭头所指为气孔;d.上表皮内表面(SEM);e.上表皮外表面(SEM);f.上表皮内表面(LM),箭头所指为气孔;g.下表皮外表面(SEM),示气孔器;h.下表皮内表面(SEM),示气孔器;i.上表皮内表面(SEM),示气孔器。 Figure 4 Epidermal characteristics of Ginkgoites aganzhenensis |
表皮构造特征:上表皮外表面较粗糙,多不平坦,密布不规则凹坑,未见乳突和毛状物(图 4d);上表皮内表面较平滑,脉络由约20行长条形细胞组成,细胞大小为(63.0~95.0) μm×(16.5~21.0) μm;脉间区细胞由多角行及四边形细胞组成,细胞大小为(32.5~37.6) μm×(25.0~32.5) μm (图 4d,f);垂周壁较直或稍弯,具不均匀加厚(图 4d);下表皮外表面多不平坦(图 4b),内表面较光滑,细胞形态同上表皮,脉络与脉间区的区别明显(图 4a)。叶片气孔式为双生型(图 4c,f),少量分布于上表皮,呈不规则排列(图 4f),下表皮气孔较多,条带状分布于脉间区,密度约为每平方毫米20个(图 4a,b,c)。气孔器为多边形、椭圆形,单环式,大小为(23.6~33.8) μm×(31.8~44.2) μm,保卫细胞肾形,略下陷;气孔孔缝长30~42 μm (图 4h,i),无定向排列;副卫细胞5~6个,近孔缝处角质增厚为乳头状突起,部分遮挡孔缝(图 4g)。
产地与层位:宝积山盆地中侏罗统窑街组。
4 讨论与比较Ginkgoites是由Seward于1919年创立的形态属,自Florin[32]于1936年对本属进行了更进一步的补充研究后,似银杏这一属名才为大多数的古植物学家所接受。Ginkgoites和Baiera均为形态属,二者在宏观形态、微细构造特征上的区别较小。通常,分裂较浅、较少的叶片一般归于Ginkgoites;分裂较深、较多的叶片归于Baiera,且Ginkgoites叶脉更加明显[33]。
杨恕等[34]于1988年首次报道了甘肃阿干镇煤田下侏罗统大西沟组的Ginkgoites一新种,根据其叶高10.5 cm,宽12 cm;4~8个裂片,梭形,长10.5 cm,宽2 cm,基部狭缩且彼此分离;表皮细胞为长条形或多边形;气孔式为双生型,保卫细胞下陷,副卫细胞6个,近孔缝处角质增厚为乳突等特征将其定名为Ginkgoites aganzhenensis Yang。与之相比,当前化石除了长度与裂片宽度有少许变化外,其他特征都非常接近,如叶片形状、叶片分裂形态和分裂次数等特征,故归入Ginkgoites确定为Ginkgoites aganzhenensis是适宜的。当前标本个体很大,裂片长达10余厘米,从这一点看,Ginkgoites magnifolius Du Toit可与之相比[35],但后者裂片数目较少,仅有4个左右,且其裂片顶端钝圆,裂片基部无当前标本这样狭缩且彼此分离的特征。Genkina[36]所描述的伊塞克-库尔盆地Ginkgoites ferganensis Brick叶片也较大,形态与本种可做比较,但这个种的裂片基部与当前标本相比无基部狭缩且彼此分离的情况,且裂片数目也较当前标本少;上述两个种均无微细构造特征描述,无法作进一步的对比。何德长等[37]于陕西北部发现的Ginkgoites gigantean He个体也较大,不同的是这个种裂片较窄,数目为6;其上表皮细胞两端尖缩,气孔孔缝也较当前标本短。厉宝贤[38]采自辽西上侏罗统的Ginkgoites truncates Li的裂片数与当前标本接近,但其裂片长度与当前标本具有明显的差别,仅有4~4.5 cm,其叶脉密度也较当前标本大。同样具有相当裂片数的还有产于英国的Ginkgo longifolius (Phillips) Harris[39],但其裂片长度与裂片宽度都较当前标本小;其叶片形态为棒状,也与当前标本具有明显差别。当前标本裂片基部狭缩且彼此分离,与Ginkgoites lepidus (Heer) Florin相似[40],但该种裂片呈线型,明显地较当前化石的窄,且拥有更多的裂片数。Harris所定东格陵兰的Ginkgoites hermelini (Hartz) Harris与当前的标本可以比较[41],但该种裂片形状以线形者为主,其裂片基部又不似当前标本那样狭缩;其上表皮气孔器数目变化很大,从极少到与下表皮近相等,气孔器侧副卫细胞角质突起多为中空,这些特征都区别于当前标本。虽然部分似银杏属和银杏属化石与当前化石在宏观形态以及微细构造上有相似性,但仍有区别(表 1)。通过对比,将当前化石定为Ginkgoites aganzhenensis是合理的。
| 种名 | 叶长/cm | 叶宽/cm | 裂片宽/mm | 裂片数/个 | 裂片形状 | 气孔式 | 气孔器排列 | 叶脉/cm |
| G. ferganensis Brick[36] | 11 | - | 16~22 | 4 | 披针形或舌形 | - | - | 6~8 |
| G. gigantean He[37] | 11 | - | 15 | 6 | 披针形 | 双生型 | 带状 | 10 |
| G. truncates Li[[38]] | 4~4.5 | 8 | - | 4~8 | 楔形 | 下生型 | 不规则 | 18 |
| G. lepidus (Heer) Florin[[40]] | 3~8 | - | 8~13 | 12~16 | 线形 | - | - | - |
| G. hermelini (Hartz) Harris[[41]] | 6~7 | 12 | 5~6 | 6~8 | 线形 | 双生型 | - | 10 |
| Ginkgo longifolius (Phillips) Harris[[39]] | 6~7 | 12 | 3.5 | 4~8 | 棒状 | 双生型 | 带状 | 15 |
| 当前化石 | 10.8 | 13 | 18 | 4~8 | 梭形 | 双生型 | 带状 | 10 |
对采自甘肃宝积山盆地中侏罗统的Ginkgoites aganzhenensis标本具气孔器的下表皮样品进行气孔参数统计,结果见表 2。通过对角质层数目与气孔参数稳定性之间的关系的研究,解三平等[42]认为,对植物化石进行气孔参数统计时,获得稳定可信的值至少需要分析4~5个角质层样本。本次研究所统计的角质层达到了8个,满足了角质层数目的可信度要求。
| 序号 | SD/mm2 | ED/mm2 | SI/% | pCO2(RF)/×10-6 | SR | pCO2(NLR)/×10-6 |
| 1 | 25 | 537 | 4.45 | 369 | 2.55 | 1 528 |
| 2 | 17 | 427 | 3.83 | 983 | 2.96 | 1 775 |
| 3 | 19 | 583 | 3.16 | 1 342 | 3.59 | 2 154 |
| 4 | 19 | 503 | 3.64 | 1 105 | 3.11 | 1 868 |
| 5 | 20 | 531 | 3.63 | 1 111 | 3.12 | 1 873 |
| 6 | 22 | 505 | 4.17 | 693 | 2.71 | 1 628 |
| 7 | 18 | 511 | 3.40 | 1 232 | 3.33 | 1 998 |
| 8 | 20 | 498 | 3.86 | 960 | 2.93 | 1 761 |
| 平均值 | 20±3 | 511±44 | 3.77 | 974±312 | 3.04 | 1 823±198 |
Ginkgoites的最近现生亲缘种(NLR)是现生银杏Ginkgo biloba L.,其气孔指数平均值为11.33%[17]。气孔参数统计结果表明宝积山盆地中侏罗统的标本SI平均值为3.77%(表 2)。在此基础上,通过气孔指数法和气孔比率法重建Ginkgoites aganzhenensis所记录的宝积山盆地中侏罗世早期的古大气CO2浓度(表 2)。
结果表明,通过气孔比率法,利用石炭纪标准得到的大气CO2浓度为1 823×10-6,而通过气孔指数法得到的大气CO2浓度为974×10-6,后者仅为前者的二分之一,且其波动也更大(表 2)。为了验证我们所获得的大气CO2浓度值的可靠性,将其投到Berner等[43]的碳平衡模型GEOCARB Ⅲ中进行对比(图 5)。

|
| 图 5 利用植物化石重建的中侏罗世的大气CO2浓度的比较及GEOCARB Ⅲ模型 Figure 5 Comparison between paleo-CO2 level of the Middle Jurassic according to fossil plants and GEOCARB Ⅲ model |
从图 5中可以看到,基于气孔指数法所得到的古大气CO2浓度值在GEOCARB Ⅲ模型可信误差范围外,气孔比率法得到的数据则在可信误差范围内,说明当使用阿干镇似银杏重建古大气CO2浓度时,气孔比率法比气孔指数法更加合适。
气孔指数法所使用的线性回归函数(RF)是在与现在的大气环境接近的CO2浓度情况下建立的,气孔指数与大气CO2浓度之间保持着良好的线性关系,但是随着大气CO2浓度的升高,存在着一个“上限”(350×10-6)[17],超过这个“上限”,则是非线性的,气孔参数对于大气CO2浓度的变化也不再灵敏[44]。因此,当大气CO2浓度低于此“上限”值时,气孔指数法所获得的结果准确度较高。侏罗纪时期古大气CO2浓度高于此界限值,因此用气孔比率法推算当前化石生长时期的古大气CO2浓度则更加准确。此外,当前化石气孔式为双生型,而使用气孔双生型的植物化石通过线性回归重建古大气CO2浓度是受到限制的[45]。利用气孔比率法可以很好地解决线性回归的缺陷[8],并能对中生代大气CO2浓度变化进行重建。
在此之前已有学者利用茨康类植物对我国部分地区中侏罗世古大气CO2浓度进行定量重建[46-47],结果证明所选用的甘肃窑街盆地中侏罗世的似管状叶属和甘肃华亭地区中侏罗世的拟刺葵属化石对大气CO2变化比较敏感。因此,茨康类植物也可以作为恢复侏罗纪大气CO2浓度的良好材料。但图 5显示,虽然利用茨康类植物与利用银杏类植物所获得的古大气CO2浓度都处于GEOCARB Ⅲ值的误差范围以内,但相较于利用同一时期银杏类植物所重建的古大气CO2浓度距GEOCARB Ⅲ最佳拟合线较远。茨康类植物现已全部灭绝,故现生Ginkgo biloba L.通常被指定为茨康类植物的现存对应种,其亲缘关系较远;而似银杏属植物与现生银杏亲缘关系较近。因此,当使用Ginkgo biloba L.作为现存对应种重建古大气CO2浓度时,似银杏属植物化石获得的结果更加准确。
5.2 古气候的重建大气中的CO2是一种非常重要的温室气体,其浓度的升高被普遍认为是温室效应的主要原因[5, 14]。甘肃宝积山盆地中侏罗世的古大气CO2浓度为1 823×10-6,可与前人在甘肃窑街盆地[48]及新疆吐哈盆地[49]利用植物化石所获得的数据相比较(图 5)。通过假设全球CO2与温度之间存在一定的函数关系,McElwain et al.[14]得到了一个关于CO2与温度的经验公式:
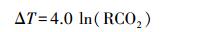 (4)
(4)
其中ΔT表示地球表面平均温度的差值。
通过公式(4),我们可以计算出当时与当今的温度差;计算结果表明温度增量为7.2℃,白银市平川区现在的温度背景值为8.9℃,则宝积山盆地在中侏罗世早期的平均温度为16.1℃,处于一种高CO2浓度、气候温暖湿润的环境中。
前文已经提到,宝积山盆地中侏罗统窑街组含大量植物化石,其植物组合为典型的奇丽锥叶蕨-似银杏组合(Coniopteris spectabilis-Ginkgoites)[27];其中:银杏类占优势,真蕨类次之,松柏类和有节类不多,苏铁类也较少。银杏类主要为似银杏属和楔拜拉属,多与茨康类伴生,基本上都是叶片化石。中生代银杏类植物十分繁盛,在世界范围内其主要分布于温带的西伯利亚-加拿大区,在中国则主要见于北方地区,南方地区极其少见,表明其主要生活在较潮湿且四季分明的气候环境中。中生代真蕨类植物种类繁多,不同的类别分别对应着不同的气候环境,但主要为喜热而潮湿的环境[50]。茨康类的茨康叶属和拟刺葵属也是中生代温带或暖温带落叶植被中的重要组成分子。这些都反映了宝积山盆地在中侏罗世早期处于温暖潮湿的温带气候环境,且植被发育良好,该结果与前人通过孢粉组合推断出的邻区窑街盆地中侏罗世早期的古气候[51-52]相互印证,这与目前宝积山盆地相对寒冷干燥、缺少植被覆盖的自然环境有很大差距。
6 结论(1)报道了采自甘肃宝积山盆地中侏罗世窑街组的Ginkgoites植物化石,通过宏观和微观特征的对比将其鉴定为Ginkgoites aganzhenensis Yang,并使用气孔指数法和气孔比率法分别定量重建了宝积山盆地中侏罗世早期的大气CO2浓度。通过与Berner等的碳平衡模型GEOCARB Ⅲ进行对比,发现通过气孔指数法得到数值处于该模型可信误差范围之外,而通过气孔比率法获得的结果则落在了可信误差范围内。表明使用Ginkgoites aganzhenensis重建古大气CO2浓度时,气孔比率法更为准确。
(2)与前人利用茨康类植物化石所获得的古大气CO2浓度值进行比较,发现通过似银杏属植物化石所获得的数据更接近GEOCARB Ⅲ最佳拟合线,表明利用当前化石获得的CO2浓度值更加准确。
(3)计算表明宝积山盆地中侏罗世早期的温度比现今高了约7.2℃,结合伴生化石组合,反映了研究区当时温暖潮湿的温带气候环境,与前人在邻区窑街盆地通过孢粉组合获得的结果可相互印证。
致谢: 感谢中国科学院兰州化学物理研究所赵家政研究员在电镜观察过程中给予的帮助。感谢兰州大学王雪莲硕士在实验过程中给予的帮助。感谢兰州大学何雨栗博士与杨国林博士在图版制作过程中给予的帮助。| [1] | Royer D L, Berner R A, Park J. Climate sensitivity constrained by CO2 concentrations over the past 420 million years[J]. Nature, 2007, 446(7135): 530–532. DOI: 10.1038/nature05699 |
| [2] | Berner R A. The rise of plants and their effect on weathering and atmospheric CO2[J]. Science, 1997, 276(5312): 544–546. DOI: 10.1126/science.276.5312.544 |
| [3] | Woodward F I. Stomatal numbers are sensitive to increases in CO2 from pre-industrial levels[J]. Nature, 1987, 327(6123): 617–618. DOI: 10.1038/327617a0 |
| [4] | Beerling D J, Royer D L. Fossil plants as indicators of the Phanerozoic global carbon cycle[J]. Annual Review of Earth and Planetary Sciences, 2002, 30(1): 527–556. DOI: 10.1146/annurev.earth.30.091201.141413 |
| [5] | Royer D L. Linkages between CO2, climate, and evolution in deep time[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(2): 407–408. DOI: 10.1073/pnas.0710915105 |
| [6] | Van Der Burgh J, Visscher H, Dilcher D L, et al. Paleoatmospheric signatures in Neogene fossil leaves[J]. Science, 1993, 260(5115): 1788–1790. DOI: 10.1126/science.260.5115.1788 |
| [7] | Royer D L, Wing S L, Beerling D J, et al. Paleobotanical evidence for near present-day levels of atmospheric CO2 during part of the Tertiary[J]. Science, 2001, 292(5525): 2310–2313. DOI: 10.1126/science.292.5525.2310 |
| [8] | McElwain J C, Chaloner W G. Stomatal density and index of fossil plants track atmospheric carbon dioxide in the Palaeozoic[J]. Annals of Botany, 1995, 76(4): 389–395. DOI: 10.1006/anbo.1995.1112 |
| [9] | Haworth M, Hesselbo S P, McElwain J C, et al. Mid-cretaceous pCO2 based on stomata of the extinct conifer Pseudofrenelopsis(Cheirolepidiaceae)[J]. Geology, 2005, 33(9): 749–752. DOI: 10.1130/G21736.1 |
| [10] | Quan Cheng, Sun Chunlin, Sun Yuewu, et al. High resolution estimates of paleo-CO2 levels through the Campanian (Late Cretaceous) based on Ginkgo cuticles[J]. Cretaceous Research, 2009, 30(2): 424–428. DOI: 10.1016/j.cretres.2008.08.004 |
| [11] | Zhou Zhiyan, Zheng Shaolin. Palaeobiology:the missing link in Ginkgo evolution[J]. Nature, 2003, 423(6942): 821–822. DOI: 10.1038/423821a |
| [12] | Sun Bainian, Dilcher D L, Beerling D J, et al. Variation in Ginkgo biloba L. leaf characters across a climatic gradient in China[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(12): 7141–7146. DOI: 10.1073/pnas.1232419100 |
| [13] | McElwain J C, Chaloner W G. The fossil cuticle as a skeletal record of environmental change[J]. Palaios, 1996, 11(4): 376–388. DOI: 10.2307/3515247 |
| [14] | McElwain J C, Beerling D J, Woodward F I. Fossil plants and global warming at the Triassic-Jurassic boundary[J]. Science, 1999, 285(5432): 1386–1390. DOI: 10.1126/science.285.5432.1386 |
| [15] | Chen Liqun, Li Chengsen, Chaloner W G, et al. Assessing the potential for the stomatal characters of extant and fossil Ginkgo leaves to signal atmospheric CO2 change[J]. American Journal of Botany, 2001, 88(7): 1309–1315. DOI: 10.2307/3558342 |
| [16] | Retallack G J. A 300-million-year record of atmospheric carbon dioxide from fossil plant cuticles[J]. Nature, 2001, 411(6835): 287–290. DOI: 10.1038/35077041 |
| [17] | Beerling D J, Royer D L. Reading a CO2 signal from fossil stomata[J]. New Phytologist, 2002, 153(3): 387–397. DOI: 10.1046/j.0028-646X.2001.00335.x |
| [18] | Sun Bainian, Xiao Liang, Xie Sanping, et al. Quantitative analysis of paleoatmospheric CO2 level based on stomatal characters of fossil Ginkgo from Jurassic to Cretaceous in China[J]. Acta Geologica Sinica:English Edition, 2007, 81(6): 931–939. DOI: 10.1111/acgs.2007.81.issue-6 |
| [19] | Xie Sanping, Sun Bainian, Yan Defei, et al. Leaf cuticular characters of Ginkgo and implications for paleoatmospheric CO2 in the Jurassic[J]. Progress in Natural Science, 2006, 16(S1): 258–263. |
| [20] | 王秋军, 徐小慧, 金培红, 等. 基于银杏类化石Baiera furcata气孔参数定量重建中生代古大气CO2浓度变化[J]. 地质论评, 2013, 59 (6): 1035–1045. [ Wang Qiujun, Xu Xiaohui, Jin Peihong, et al. Quantitative reconstruction of Mesozoic paleoatmospheric CO2 based on stomatal parameters of fossil Baiera furcata of Ginkgophytes[J]. Geological Review, 2013, 59(6): 1035–1045. ] |
| [21] | Wang Yongdong, Huang Chengmin, Sun Bainian, et al. Paleo-CO2 variation trends and the Cretaceous greenhouse climate[J]. Earth Science Reviews, 2014, 129: 136–147. DOI: 10.1016/j.earscirev.2013.11.001 |
| [22] | 王永栋, 孙柏年, 黄成敏, 等. 地史时期古大气二氧化碳变化趋势与温室气候--以中生代白垩纪为例[J]. 自然杂志, 2015, 37 (2): 108–114. [ Wang Yongdong, Sun Bainian, Huang Chengmin, et al. Variation of paleo-CO2 and greenhouse climate in the geological history:A case study from the Cretaceous of the Mesozoic[J]. Chinese Journal of Nature, 2015, 37(2): 108–114. ] |
| [23] | 王佟.中国西北赋煤区构造发育规律及构造控煤研究[D].北京:中国矿业大学(北京), 2012. [ Wang Tong. Tectonic development law and tectonic coal-controlling of northwest China coal hosting area[D]. Beijing:China University of Mining & Technology, Beijing, 2012. ] |
| [24] | 张泓, 何宗莲, 晋香兰, 等. 祁连山东段宝积山-红会煤盆地沉积环境与聚煤规律[J]. 沉积学报, 2009, 27 (4): 622–631. [ Zhang Hong, He Zonglian, Jin Xianglan, et al. Sedimentary environments and coal accumulation of the Baojishan-Honghui Basin, eastern Qilian Mountains[J]. Acta Sedimentologica Sinica, 2009, 27(4): 622–631. ] |
| [25] | 徐亚军, 杜远生, 杨江海, 等. 甘肃靖远上泥盆统沙流水组沉积地球化学特征及其物源分析[J]. 沉积学报, 2011, 29 (1): 41–54. [ Xu Yajun, Du Yuansheng, Yang Jianghai, et al. Geochemistry and provenance of the Upper Devonian Shaliushui Formation in Jingyuan county, Gansu province[J]. Acta Sedimentologica Sinica, 2011, 29(1): 41–54. ] |
| [26] | 李菊花, 谭志伟, 关保国. 靖远煤田宝积山-红会矿区外围赋煤特征及勘查方向研究[J]. 中国煤炭地质, 2012, 24 (7): 7–10. [ Li Juhua, Tan Zhiwei, Guan Baoguo. A study on coal hosting features and prospecting orientation in peripheral Baojishan and Honghui Mine areas, Jingyuan coalfield[J]. Coal Geology of China, 2012, 24(7): 7–10. ] |
| [27] | 孙柏年. 甘肃靖远煤田早中生代植物化石组合序列[J]. 兰州大学学报:自然科学版, 1992, 28 (3): 163–167. [ Sun Bainian. The assemblage succession of the early Mesozoic fossil plants from Jingyuan coal field in Gansu[J]. Journal of Lanzhou University:Natural Sciences, 1992, 28(3): 163–167. ] |
| [28] | 刘子进. 甘肃东部早侏罗世地层及植物群的初步研究[J]. 中国地质科学院西安地质矿产研究所所刊, 1982 (5): 88–100. [ Liu Zijin. A preliminary study of early Jurassic beds and the flora in eastern Gansu[J]. Bulletin of Xi'an Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences, 1982(5): 88–100. ] |
| [29] | 倪庆, 王永栋, 田宁, 等. 从植物化石叶片中获取古大气CO2浓度的信息[J]. 自然杂志, 2008, 30 (2): 102–105. [ Ni Qing, Wang Yongdong, Tian Ning, et al. Tracking palaeo-CO2 information from fossil plant leaves[J]. Chinese Journal of Nature, 2008, 30(2): 102–105. ] |
| [30] | McElwain J C, Mayle F E, Beerling D J. Stomatal evidence for a decline in atmospheric CO2 concentration during the Younger Dryas stadial:a comparison with Antarctic ice core records[J]. Journal of Quaternary Science, 2002, 17(1): 21–29. DOI: 10.1002/(ISSN)1099-1417 |
| [31] | McElwain J C. Climate-independent paleoaltimetry using stomatal density in fossil leaves as a proxy for CO2 partial pressure[J]. Geology, 2004, 32(12): 1017–1020. DOI: 10.1130/G20915.1 |
| [32] | Florin R. Die fossilen Ginkgophyten von Franz-Joseph-Land nebst Erörterungen über vermeintliche Cordaitales mesozoischen alters. I. Spezieller tei[J]. Palaeontographica Abteilung B, 1936, 81: 71–173. |
| [33] | 斯行健, 李星学. 中国植物化石(二):中国中生代植物[M]. 北京: 科学出版社, 1963: 1-429. [ Si Xingjian, Li Xingxue. Mesozoic Plants from China:Fossil Plants of China Ⅱ[M]. Beijing: Science Press, 1963: 1-429. ] |
| [34] | 杨恕, 孙柏年, 沈光隆. 兰州地区侏罗纪似银杏属的新材料[J]. 兰州大学学报:自然科学版, 1988, 24 (S1): 70–77. [ Yang Shu, Sun Bainian, Shen Guanglong. New materials of Ginkgoites from Jurassic in vicinity of Lanzhou, Gansu[J]. Journal of Lanzhou University:Natural Sciences, 1988, 24(S1): 70–77. ] |
| [35] | Du Toit A L. The Fossil Flora of the Upper Karroo Beds[M]. London: Trustees of the South African Museum, and the Geological Survey of South Africa, 1927: 289-420. |
| [36] | Genkina R Z. Fossil Flora and Stratigraphy from Older Mesozoic Strata of Issyk-kul Basin[M]. Moscow: Nauka, 1966: 1-148. |
| [37] | 钱丽君, 白清昭, 熊存卫, 等. 陕西北部侏罗纪含煤地层及聚煤特征[M]. 西安: 西北大学出版社, 1987: 81-82. [ Qian Lijun, Bai Qingzhao, Xiong Cunwei, et al. Jurassic Coal-Bearing Strata and the Characteristics of Coal Accumulation from Northern Shannxi[M]. Xi'an: Northwest University Press, 1987: 81-82. ] |
| [38] | 厉宝贤. 辽西阜新海州组的四种似银杏化石的表皮构造[J]. 古生物学报, 1981, 20 (3): 208–215. [ Li Baoxian. On the cuticular structures of Ginkgoites from the Haizhou Formation (Upper Jurassic) in Fuxin (Fushin), western Liaoning[J]. Acta Palaeontologica Sinica, 1981, 20(3): 208–215. ] |
| [39] | Harris T M, Millington W, Miller J. The Yorkshire Jurassic Flora, IV[M]. London: British Museum (Natural History), 1974: 21-26. |
| [40] | Von Heer O. Beiträge zur Jura-flora Ostsibiriens und des Amurlandes[J]. Mémoires de l'Académie Impériale des Sciences de Saint Pétersbourg, 1876, 22(12): 1–122. |
| [41] | Harris T M. The fossil flora of Scoresby Sound, East Greenland, Part 4:Ginkgoales, Coniferales, Lycopodiales and isolated fructifications[J]. Meddelelser om Grönland, 1935, 112(1): 1–176. |
| [42] | 解三平, 阎德飞, 韦利杰, 等. 精确重建古大气CO2浓度的气孔方法[J]. 古生物学报, 2005, 44 (3): 464–471. [ Xie Sanping, Yan Defei, Wei Lijie, et al. A stomatal approach for accurate reconstruction of palaeoatmospheric CO2 concentration[J]. Acta Palaeontologica Sinica, 2005, 44(3): 464–471. ] |
| [43] | Berner R A, Kothavala Z. GEOCARB Ⅲ:a revised model of atmospheric CO2 over Phanerozoic time[J]. American Journal of Science, 2001, 301(2): 182–204. DOI: 10.2475/ajs.301.2.182 |
| [44] | Royer D L, Hickey L J, Wing S L. Ecological conservatism in the "living fossil" Ginkgo[J]. Paleobiology, 2003, 29(1): 84–104. DOI: 10.1666/0094-8373(2003)029<0084:ECITLF>2.0.CO;2 |
| [45] | Royer D L. Estimating latest Cretaceous and Tertiary atmospheric CO2 from stomatal indices[J]. Geological Society of America Special Papers, 2003, 369: 79–93. |
| [46] | 闫德飞, 孙柏年. Solenites murrayana L. et H.在甘肃窑街煤田的发现及其地质意义[J]. 兰州大学学报:自然科学版, 2004, 40 (3): 84–88. [ Yan Defei, Sun Bainian. The discovery of Solenites murrayana L. et H. in Yaojie coal field, Gansu and its geological significance[J]. Journal of Lanzhou University:Natural Sciences, 2004, 40(3): 84–88. ] |
| [47] | 丁素婷, 孙柏年, 吴靖宇, 等. 甘肃华亭侏罗系Phoenicopsis (Phoenicopsis) angustifolia Heer的表皮构造与碳同位素特征[J]. 兰州大学学报:自然科学版, 2010, 46 (1): 14–21. [ Ding Suting, Sun Bainian, Wu Jingyu, et al. Cuticular composition and carbon isotope characteristics of Jurassic Phoenicopsis (Phoenicopsis) angustifolia Heer from Huating in Gansu province, China[J]. Journal of Lanzhou University:Natural Sciences, 2010, 46(1): 14–21. ] |
| [48] | Yan Defei, Sun Bainian, Xie Sanping, et al. Response to paleoatmospheric CO2 concentration of Solenites vimineus (Phillips) Harris (Ginkgophyta) from the Middle Jurassic of the Yaojie Basin, Gansu Province, China[J]. Science in China Series D:Earth Sciences, 2009, 52(12): 2029–2039. DOI: 10.1007/s11430-009-0181-1 |
| [49] | 肖良, 李勇, 周建仁, 等. 新疆吐哈盆地中侏罗世古大气CO2重建[J]. 兰州大学学报:自然科学版, 2014, 50 (2): 154–160. [ Xiao Liang, Li Yong, Zhou Jianren, et al. Paleoatmospheric CO2 level of the Middle Jurassic in Turpan-Hami Basin, Xinjiang[J]. Journal of Lanzhou University:Natural Sciences, 2014, 50(2): 154–160. ] |
| [50] | 邓胜徽. 中生代主要植物化石的古气候指示意义[J]. 古地理学报, 2007, 9 (6): 559–574. [ Deng Shenghui. Palaeoclimatic implications of main fossil plants of the Mesozoic[J]. Journal of Palaeogeography, 2007, 9(6): 559–574. ] |
| [51] | 辛存林, 孙柏年, 韦利杰, 等. 甘肃窑街中侏罗世孢粉组合及其古环境意义[J]. 兰州大学学报:自然科学版, 2006, 42 (3): 8–13. [ Xin Cunlin, Sun Bainian, Wei Lijie, et al. Sporo-pollen assemblage of the Middle Jurassic period from Yaojie, Gansu and its paleoenvironmental significance[J]. Journal of Lanzhou University:Natural Sciences, 2006, 42(3): 8–13. ] |
| [52] | 辛存林, 陈世伟, 都卫东, 等. 甘肃窑街中侏罗世Coniopteris margaretae的原位孢子与分散孢子的关系[J]. 地质科技情报, 2011, 30 (5): 36–40. [ Xin Cunlin, Chen Shiwei, Du Weidong, et al. Relationship between spore in situ and dispersed spore of Coniopteris margaretae of the Middle Jurassic from Yaojie, Gansu province[J]. Geological Science and Technology Information, 2011, 30(5): 36–40. ] |