 2017, Vol. 35
2017, Vol. 35扩展功能
文章信息
- 胡建芳, 彭平安
- HU JianFang, PENG PingAn
- 有机地球化学研究新进展与展望
- An Overview and Perspectives on Organic Geochemistry
- 沉积学报, 2017, 35(5): 968-980
- ACTA SEDIMENTOLOGICA SINCA, 2017, 35(5): 968-980
- 10.14027/j.cnki.cjxb.2017.05.009
-
文章历史
- 收稿日期:2017-01-03
- 收修改稿日期: 2017-03-25



沉积岩/物是地质历史时期环境—生物协同演化过程的重要记录者。沉积地球化学综合地质、沉积和古生物记录,并运用数值模拟等手段,重建元素循环及地质历史演化。其中的有机地球化学是研究沉积体有机质的分布、迁移、富集与转化机制的地球化学分支学科。生物死亡后的有机质演化及其地球化学过程都属于这一学科的研究范畴。在国际上,其研究大致可以分成三个领域:石油地球化学、环境有机地球化学和生物—有机地球化学。在这里,我们把中国科学院学部学科发展战略研究——“沉积学发展战略研究”项目中取得的相关认识做一梳理,着重就近五年来有机地球化学研究的新进展进行分析和总结,并就新世纪我国有机地球化学研究的增长点做一展望。
1 国际有机地球化学研究的主要进展近年来,随着各种谱学、化学方法的发展和仪器检测水平的提高,国际有机地球化学的研究取得了一系列重要的进展。这些进展包括:1) 对沉积有机质的性质和结构的认识有了较大的提高。目前,我们能在分子和分子同位素水平上了解地质体中的有机质,从而产生了大量的分子及其同位素数据,大大促进了有机地球化学的发展。2) 对有机质参与的地球化学全过程能够进行定性的研究。在水柱阶段,了解了各种微生物对不同有机质的改造降解规律[1-2];在成岩作用阶段,已可定量化表征有机质的热演化过程[3-5];在后生作用阶段,基本上弄清了一些主要反应,如硫酸盐热化学还原反应等。3) 石油地球化学从烃源岩的研究走向了成藏研究。通过成藏记录理清成藏过程在烃源岩、疏导层、储层等节点上的表现,从而揭示油气藏的形成规律。4) 环境有机地球化学逐步发展壮大。环境有机地球化学在污染控制领域发挥着重要作用,特别是在二次污染的源解析、污染过程的研究方面作出了其他学科不能替代的贡献。5) 生物—有机地球化学蓬勃发展。随着新技术、新方法的发展,生物—有机地球化学在稳定碳的形成、定量环境变化与古生态系统重建研究中发挥了举足轻重的作用。对于前四个方面的进展,国内的有机地球化学家已有不少总结,我们在这里不再赘述。生物—有机地球化学是近几年国际有机地球化学研究的热点,进展较快且成果较多,有必要花些笔墨重点介绍。
1.1 海洋生态环境研究 1.1.1 表层海水温度上个世纪80年代,基于C37长链不饱和烯酮不饱和度的U37K温标的诞生是有机地球化学对海洋生态环境研究的创新性贡献[6]。C37长链烯酮是一类由定鞭金藻纲特别是颗石藻合成的含两个至四个C=C双键的特征生物标志化合物,在全球大洋水体和沉积物中广泛存在[7]。通过现场采样、培养实验和大洋表层沉积物的系统分析,都揭示出U37K与定鞭金藻生长的水体温度存在良好的线性关系(其中,U37K= 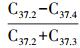
TEX86(TetraEtherindeX of tetraethersconsisting of 86 carbon atoms)指标是近20年来发展起来的另一类表层海水温度替代指标[9]。近五年来,国际上发表的与TEX86相关的论文呈指数形式增长。但随着研究的进行,发现TEX86反映的是海洋次表层温度,而不是表层温度,且影响TEX86的因素不仅仅是温度[10-11]。因此,在古环境研究中,通常需要多指标的相互校验,这不仅有助于获得相对准确的古环境数据,而且有可能从各指标的差异中了解潜藏的环境与生物相互作用的机理。
近几年发展起来的另一个用来计算SST的有机地球化学指标是长链二醇指数(Long chain Diol Index,LDI = 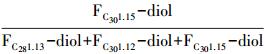
近年来,碳循环问题日益成为全球变化与地球科学研究领域的前沿与热点问题。碳是主要的生源要素之一,是有机质的重要组成部分。地球上主要有四大碳库,即大气碳库、海洋碳库、陆地生态系统碳库和岩石圈碳库。其中海洋碳库的碳是大气碳库的60多倍[18],在全球碳循环中起着十分重要的作用。从千年尺度上看,海洋浮游生物泵在或者说海洋初级生产力的变化对大气CO2浓度起了重要的调节作用(图 1)[19-20]。海洋初级生产力和浮游植物生态群落结构的改变通过影响有机泵和碳酸盐泵的组成而直接影响海洋碳埋藏效率。如海洋浮游植物种群结构从以颗石藻为主向以甲藻或硅藻为主转变时,海洋CaCO3/Corganic“沉积雨”比值(rain ratio)会下降,海洋水体pH值会升高,最终会引起大气CO2的降低。

|
| 图 1 海洋中生物泵简化示意图 Figure 1 Simplified diagram showing the biological pump in the ocean |
全球海洋古生产力的定量估算有多种方法,其中利用沉积有机碳含量计算输出生产力是最经典的方法。在利用沉积有机碳方法估算海洋初生产力时必须扣除陆源有机碳输入的影响,因为即使在远洋地区,陆源有机碳对沉积有机碳的贡献也不容忽视[21-22]。这时就需要通过有机地球化学的方法来估算海洋中陆源有机碳的组成比例,从而得到相对更准确的海洋初生产力数据[23]。因此,生物—有机地球化学在海洋古生产力与碳循环研究中发挥了重要作用。
生物—有机地球化学对于研究海洋初级生产力的贡献者─浮游植物种群结构的历史变化具有其他方法不可比拟的优势。如可以利用生物标志化合物反演历史时期海洋浮游植物群落结构:甲藻甾醇指示甲藻、长链烯酮指示颗石藻、菜籽甾醇指示硅藻。阿拉伯海沉积钻孔的研究就揭示出近20万年来,尽管其沉积有机碳和海洋初级生产力发生了显著变化,但甲藻甾醇、长链烯酮和菜籽甾醇之间的相对含量一直保持稳定,反映出其浮游植物种群结构并没有发生明显改变[24]。氨基酸单体化合物的稳定C同位素揭示出赤道北太平洋近千年来对于输出生产力有重要贡献的微藻种群结构的显著变化:中世纪暖期以非固氮蓝藻为优势种,到小冰期转变成以真核微藻为主,工业革命以来,则以固氮蓝细菌为优势种[25]。生物标志化合物的研究也表明巴西东南海域在末次盛冰期由于海平面的降低,陆源输入的增多使得海洋初级生产力升高,沉积有机碳也同时增加[26]。当然,由于不同生物标志化合物的差异性降解以及专属性生物标志化合物的缺乏,现有的生物标志化合物法需要同其他方法如微体古生物学、无机元素地球化学等进行综合比对才能得出更合理的认识。
1.2 陆地生态环境研究 1.2.1 古湖泊环境同海洋相比,湖泊汇水域小、沉积速率大,是区域高分辨率古环境、古气候重建研究的理想场所。湖泊沉积可完整记录地质历史时期区域的气候、植被以及人类活动的演化轨迹,且可以获得百年、甚至十年尺度的古气候事件,因此,湖泊沉积成为当前全球变化研究的焦点所在。当前的研究成果表明,湖泊沉积有机质在重建陆相古温度、植物群落演替、古大气二氧化碳分压(pCO2)等研究方面发挥着其他地质体/载体无可替代的作用。
在陆相古温度重建研究中,生物—有机地球化学发挥了绝对的优势作用。之前关于陆相古温度定量重建一直难以取得突破,是因为缺乏有效的定量化计算指标。近年来,基于支链GDGTs的陆相古温度指标得到了广泛的应用与发展。最初的研究认为支链GDGTs来源于陆地土壤和泥炭细菌,基于此,国际上建立起了土壤的年平均大气温度(MAT,Mean annual Air Temperature) MBT/CBT-MAT经验计算式[27]。但新近的研究指出支链GDGTs可以由湖泊内源产生[28],因此,出现了一些基于湖泊现代表层沉积物的MAT经验计算式[29],虽然这些计算式是“区域性”的,但其计算结果仍然具有广泛的适用性。
另一方面,被广泛运用于海洋的U37K温标近年来在湖泊中的应用也有了长足的发展。长链烯酮化合物主要在咸水湖泊中检出,而最近的研究在北美[30-31]、欧洲[32-33]以及亚洲[34]的一些淡水湖泊中都检测出了这类化合物。通过对格陵兰及北美淡水和低盐湖泊现代过程的研究,发现湖泊中长链烯酮化合物作为温度指标还存在的一些问题,一是不同类型湖泊中这类化合物生物源─定鞭藻的种属可能不同,二是不同湖泊定鞭藻生长的季节不同,这就导致U37K与温度的计算式需要进行区域和季节的校正[32, 35-37],从而使得长链烯酮在湖泊中的应用更加复杂。但随着更多生物学如DNA、18sRNA技术的应用,湖泊中长链烯酮的生源解释将更全面、真实,这类化合物在湖泊中的应用也将会更广泛。
类脂化合物特别是高碳数正构烷烃的δD值也是一个受到广泛关注的指标,因其是重建陆地水环境变化的重要手段,在湖泊和泥炭沉积物中得到了一定的应用。近年来对青藏高原湖泊中高碳数正构烷烃δD值的研究,就发现印度夏季风在早全新比较强盛,且受北半球太阳辐射影响[38-39]。但由于来源于高等植物的化合物的单体δD值受水源和其他许多诸如环境、物理/化学等因素的影响[40],常使得其δD的应用受到一定的限制。未来湖泊沉积物中来源于水生生物的化合物单体δD的应用将会受到广泛重视。
1.2.2 C3/C4植被根据光合作用固碳途径的不同,陆地高等植物可分为C3,C4和CAM三类。C3植物主要包括树木、灌木和其他喜温凉气候的草本植物,适应于湿润和较高大气二氧化碳分压(pCO2)气候条件,固碳过程中碳同位素分馏较强,其δ13C值集中在-22‰至-34‰之间(平均为-27‰)。C4植物主要包括生活在干旱热带地区的草本植物,适应于低pCO2、高温、高光照、干旱和盐碱化等环境条件,固碳过程中碳同位素分馏较弱,其δ13C值集中在-10‰至-14‰之间(平均为-13‰)。研究显示,植物合成的高碳数正构烷烃等类脂物的δ13C值比其总体δ13C值约偏负-6‰[41]。因此,沉积记录的有机碳δ13C值和来源于高等植物的类脂化合物的单体δ13C值成为重建地质历史时期陆地C3/C4植被有效手段[42-44]。
C4光合作用途径的出现是地球植物发展史上一次重要进化变革事件[45]。C4光合作用的优势在于它的CO2浓缩机制,使得核酮糖-1, 5-二磷酸羧化酶/加氧酶(Rubisco)的氧化反应达到最小化,从而尽可能避免光呼吸作用的发生。关于C4植物的起源时间虽然尚无定论,但一些研究表明C4植物早在渐新世就已出现[46],只是当时所占比例较少。自晚中新世后,C4植物经历了几次显著的扩张使其在陆地生态系统中的丰度大幅增加,但发生扩张的时间在不同地区是有差异的。C4植物扩张的原因说法不一,但目前的研究普遍认为除了气候之外,大气CO2分压、温度以及自然大火等都是影响C4植物扩张的因素[47-48]。
在利用沉积记录的总体有机碳重建地质历史时期C3/C4植被比例时,由于C3/C4植被δ13C端元值的选择不同,普遍可导致高达20%的计算误差[49],这是由于树、灌木以及开花植物总体有机碳的δ13C变化范围为-21‰至-35‰[50]。因此,来源于高等植物的特征生物标志化合物的单体δ13C值被优先用来重建地质历史时期C3/C4植被比例。但由于不同种植物或者同一植物的不同部位或不同生长期,其高碳数正构烷烃的分子组成和单体化合物的δ13C值都存在较大差异[51-52],所以在进行地质记录的重建研究时,需要结合现代过程的研究,综合考虑区域的植被类型。
1.3 与烃源岩发育相关的环境与控制因素烃源岩的发育需要大规模有机质的堆积,地质历史时期海洋大规模的有机质堆积形成了现今约80%以上的原油。这类有机质的母质主要为藻类,富含脂类物质。沉积有机质母源研究极大地促进了不同生物有机质稳定成分以及生物有机质死亡后水柱过程的研究。精确的母源与过程识别技术使我们可以从沉积有机质的研究中重建水柱的古生物地球化学过程[53],解决什么是水柱的初级生产力、进行详细的水柱分层与细菌分解过程研究等[54]。
从重建古生物地球化学过程的大量研究中,研究者发现不同时期,沉积有机质形成过程具有很大的差别,从而激发人们对沉积有机质形成机制的研究。这一工作也使不少有机地球化学家认识到,控制海洋大规模有机质堆积的原因可能是特殊的生物地球化学过程,即营养盐的特殊富集机制。但如何在地质体找到相关证据,并将其有机地联系在一起,是目前研究的难题。
高的生产力和有利的有机质保存环境是海洋大规模有机质堆积的必要条件。通常还原环境有利于有机质的保存[55],这在低生产力的情况下显得十分的重要,在高生产力条件下,由于有机质的充分供给,水体底部基本是还原的,还原环境不足以成为主要控制因素,而高的生产力成为高有机质堆积决定性因素。从目前的研究大致推测导致显生宙海洋大规模有机质堆积事件发生的营养盐的特殊富集机制不外乎三种情况。
第一种机制是火山活动提供营养物质,火山活动发生的时间较短,只要面积够大,火山灰溶解后就可提供大量的营养物质。晚泥盆/早石炭世(集中在374~345 Ma)、早白垩世(集中在125~93 Ma)的高有机质堆积可能与此有关。
第二种机制是大规模冰期后的营养物质供给。冰期存在明显的物理风化,但这些物质由于冰的固结作用,输运到海洋十分困难,在冰后期,这些营养物质在较短的时间进入海洋,可造成富营养化,从而引有机质的堆积。前寒武纪的玛丽诺与斯图特冰期、晚奥陶/早志留世(集中在445~439 Ma)冰期,极有可能为后来的大规模有机质堆积提供营养。
第三种机制是极端干旱之后的营养盐供给。干旱条件下的物理风化造成了营养物质在陆上的累积,在后期的气候转换阶段有可能大规模进入海洋,从而为藻类勃发提供条件。中国东部的古近纪湖泊中的高有机质层,有可能就是这样形成的。
上述三种情况均是极端条件下造成的特殊营养供给。对于具体的层位,可能存在多种机制并存的状况,如火山活动有可能使气候变冷,从而产生冰川,两者有可能在一起对有机质的堆积起了作用,使得烃源岩发育。这三种假设都没有得到沉积记录的验证,需要未来更多的地球化学工作去揭示和验证。
2 我国有机地球化学研究的主要进展我国有机地球化学研究起步较晚,初期完全以国内能源需求为导向,以国内特有的陆相油气资源为研究对象,发现和证实了系列湖相生物标志物,建立和完善了相关油气形成理论,在石油地球化学领域做出了具有中国特色的研究贡献。近年来,围绕大众关注的环境变化和环境污染问题,有机地球化学研究方向逐步拓展到了生物—有机地球化学和环境有机地球化学方面,也取得了长足的发展。
2.1 陆相地层的特征生物标志物陆相沉积盆地面积相对较小,水体小而窄,小盆、深盆深水沉积通常具有高的沉积速率和高频旋回,使得湖相沉积环境发生从淡水到咸水、从浅水到深水的巨大变化,形成油页岩、蒸发岩、泥炭沼泽和煤等多种沉积物;有机质的生源输入也相应发生较大变化:煤沉积以高等(木本)植物为主,泥炭沼泽沉积以草本植物为主,淡水湖相沉积以藻类为主,高盐咸水沉积以嗜盐菌藻类为主,到了蒸发岩高盐沉积环境除嗜盐菌藻类外还可能有分层水体中的光合细菌存在;沉积有机质的保存条件也可以从含氧沉积(泥炭沼泽和煤)变化到强还原沉积(高盐环境)。这些差异,决定了陆相沉积地层中生物标志物类型及其分布变化较大,不同沉积环境具有不同的特征生物标志物。
2.1.1 4, 4-二甲基甾烷从生物化学意义上看,来自法呢基焦磷酸脂(FPP)的角鲨烯变为羊毛甾醇和环阿屯醇。后者在植物中常常转化为豆甾醇,前者在动物中多见,且易转化为胆甾醇。这类生物标志物主要在水柱过程之后、沉积物埋藏压实后的地层中或者生物降解显著的油气藏中检出。在晋县凹陷赵兰庄酸性H2S油气藏的原油样品中,采用正常全扫描模式检测到了丰富的短链羊毛甾烷、4, 4-二甲基甾烷和4-甲基甾醇[56]。推断油藏早期发生了喜氧甲烷氧化菌的复苏和繁盛,这些细菌利用、消耗了地层水中的氧,使得油藏变为厌氧环境;然后厌氧甲烷氧化菌和硫酸盐还原菌发育,产生大量H2S。因为能同时产生羊毛甾醇、4, 4-二甲基甾醇和4-甲基甾醇的主要是甲烷氧化菌(Methylotroph)。文献一般认为,C27甾烷来自藻类,C29甾烷来自高等植物,4-甲基甾烷来自硅藻。在该降解油藏案例中,C1~C3甲基甾烷却来自细菌。至于其短链成因,推测可能与微生物的侧链降解断裂以及H2S对甾醇侧链上的烯键进行攻击而成的。
2.1.2 芳基类异戊二烯化合物2, 3, 4-甲基和2, 3, 6-甲基结构芳基类异戊二烯化合物分别来自光合细菌——紫硫菌和绿硫菌。它们通常处于盐湖盆地分层水体下部厌氧环境,以底部沉积底泥中硫酸盐还原菌生成的H2S气体为营养源,在生物化学作用过程中生成很多带有双键的、具有类异戊二烯结构的类胡萝卜素化合物,经成岩地球化学演化成为2, 3, 4-甲基和2, 3, 6-甲基结构的芳基类异戊二烯化合物。
柴达木盆地发育有2, 3, 4-甲基和2, 3, 6-甲基芳基类异戊二烯[57],但是江汉盆地原油中的芳基类异戊二烯化合物双峰型更为典型[58],呈现1:1的等分分布,说明水体中同时存在紫硫菌和绿硫菌,或者有绿硫菌和紫硫菌共生体“Chlorochromatium”存在。进而说明水体中光合紫硫菌和绿硫菌对烃源岩有机质的形成有重要贡献,其光合作用生成叶绿素和胡萝卜素,实质上是初始生产者;含紫硫菌和绿硫菌水体通常是一种光合细菌主导下的分层水体生态和沉积模式。这种沉积模式,对其中的特征性生物标志物的分布给出了良好的解释。双峰型芳基类异戊二烯生物标志物的发现,揭示主要是由分层水体中紫硫菌和绿硫菌的贡献造成的,位于化跃层之上嗜菌纤毛虫,以紫硫菌和绿硫菌为食,在缺少甾醇供应条件下生成伽马蜡烷的前身物四膜虫醇,是江汉盆地高丰度伽马蜡烷主要的生物源。细菌叶绿素的植基、法尼基侧链对高丰度植烷和类异戊二烯也有重要贡献[58]。这类化合物的检出对研究古湖泊生态具有重要意义。
2.1.3 其他化合物与苯并藿烷相比,25-降苯并藿烷的研究较少,Tian et al.[59]在我国川西北泥盆系的降解沥青中检测鉴定出了一系列的25-降甲基苯并藿烷、25-降甲基-2-甲基苯并藿烷,为油气源对比和环境探讨提供了重要手段。李敏等[60]在塔里木下奥陶系储层稠油中也检测到了完整系列的25-降苯并藿烷(C31~C34),且发现25-降苯并藿烷与苯并藿烷的比值与现有的生物降解评价参数有较好的可比性,推测25-降苯并藿烷系列化合物有可能作为一种研究原油生物降解的新指标。除此之处,为了研究油气的充注方向,李美俊等[61-62]新鉴定14个C3-和C4-二苯并噻吩与3个苯并萘并噻吩异构体,且论证了这些化合物运移分馏效应,建立其示踪指标体系,成功用于地层中油气充注途径研究。
2.2 煤成烃、未成熟油和深部油气煤成烃是指煤系烃源岩(煤层和煤系泥岩)有机质在成煤过程及热成熟过程中生成的石油和天然气[63]。煤成烃的研究与煤成油气的勘探密切相关[64]。上世纪五、六十年代,全球大气田和天然气储量的70%~80%来源于煤系烃源岩[64-65]。鉴于国内煤炭资源极为丰富而探明煤成气储量很低,近年来,国内油气地球化学家一直将煤成烃作为重要的研究课题。通过研究揭示至2011年底,国内煤成气总探明储量为5.8×1012 m3,占国内天然气总探明储量的69.72%[64-65]。来源于煤系烃源岩原油的储量远低于煤成气的储量,主要分布在吐鲁番—哈密盆地、准噶尔盆地、焉耆盆地和塔里木盆地库车坳陷等。
未成熟油是在成岩作用晚期,沉积岩中有机质的成烃演化达到生油门限之前所形成的石油。其烃源岩成熟度范围约在镜质体反射率0.3%~0.7%之间,其原油性质一般为重质石油,也有凝析油。未成熟油的生烃母质及成因机理:1) 非烃、沥青质生成的高含硫原油;2) 脂肪酸成烃;3) 树脂体成烃;4) 干酪根早期降解成烃;5) 藻类生物类脂物的早期成烃;6) 生物作用早期成烃;7) 木栓质体早期成烃。国内在济阳坳陷、江汉盆地、苏北盆地、渤海湾盆地、南阳盆地、柴达木盆地和百色盆地等盆地都发现了低成熟原油。
深部油气具有三个方面的含义:1) 东部盆地埋深>3 500 m[66],西部盆地埋深>4 000 m的油气藏[67];2) 成熟度很高(过成熟)的气藏;3) 与叠合盆地古生界海相烃源岩相关的油气藏。近十年来,在四川盆地发现了包括普光和川中特大气田在内的一大批大气田,这些气田的含气层位埋深均大于4 000 m,气源主要来自古生界海相烃源岩,烃气组成以甲烷为主,湿气含量很低,C1/C1-4比值介于0.95~1.0之间[68]。经过多年的勘探工作,认为深部油气已成为我国主要的勘探方向和目标[69-73]。
2.3 古生态、古环境重建由于生物标志化合物及其碳氢氮等同位素比值是很好的古生态、碳循环、水循环和氮循环等的示踪物或代用指标,有机地球化学在地球表层系统环境演变研究中得到越来越广泛的重视和应用。我国学者在这一领域的研究工作也逐渐接近国际前沿。这一领域的工作主要集中于如下两大方面:1) 代用指标的现代过程与适用性研究;2) 运用代用指标进行古生态、古环境的历史重建。
在代用指标的现代过程与适用性研究上,Sun et al.[74]通过对不同纬度带数十个不同类型湖泊沉积物中长链烯酮的分布,系统总结了各类湖泊体系中U37K温标的经验计算式的优缺点,重新建立了U37K’温标与平均水温的经验计算式。Song et al.[75]通过调查中国西北近20个湖泊中长链烯酮的分布,发现U37K温标受盐度的影响显著,而U37K’受盐度影响有限,更适合于中国西北湖泊水体温度的重建。此外,对由奇古菌(Thaumarchaeota)和未知细菌的甘油二烷基甘油四醚类化合物(GDGT)构建的TEX86和CBT/MBT温标,我国学者也开展了广泛和深入的研究,对这些指标在我国湖泊、干旱区土壤和边缘海区的适用性和指示意义都分别进行了详细探讨[76-81]。
但U37K’温标重建湖泊温度的工作在我国的应用尚不多见[82-85]。而U37K温标在我国南海和东黄海不同时间尺度的古温度重建方面都得到了成功的应用[86-89],为研究西太平洋暖池始新世以来的历史、冰期旋回中季风演变历史等方面的研究提供了关键数据。运用与GDGTs相关的指标重建古环境方面发表的成果还相对较少[90-94],可能与上述指标在国内传统古环境古气候实验室尚未得到普及性应用有关。
有机地球化学在地质历史古生态重建研究中也发挥了重要的作用。古生态涉及的生物类型主要有陆地高等植物、水生浮游植物、微生物三大类。1) 来源于高等植物的黑碳和叶蜡类脂单体的碳同位素可以探索陆地植被中C4植物的起源和C3/C4植物的比例,进而反映古气候的演变历史。我国学者从海洋、黄土和湖泊沉积物中都针对上述问题进行了不同时间尺度的研究工作[95-97];2) 运用海洋沉积中的甾醇类、长链烯酮以及二醇类化合物等可以重建海洋浮游植物的种群结构变化,进而了解海洋生物泵和营养盐变化的历史。这方面的工作在我国边缘海古海洋研究中得到了应用[98-99]。3) 微生物类型丰富、数量巨大,是联系其他生物和环境的重要纽带,在碳、氮、硫和金属元素循环中发挥着重要作用。与之相关的生物标志物类型丰富而众多。我国学者在这方面的突出工作是对浙江长兴煤山Tr/P界线附近的微生物(蓝细菌、硫细菌等)分子记录的深入研究,揭示了这一重大地质事件过程中环境的不稳定性和生物危机的多阶段性[100-101]。
水循环历史重建是古气候重建的关键内容。生物标志物分子中氢元素的初始来源为生物所利用的环境水,因此氢同位素(δD)反映了源水同位素特征,且其变化受气候和环境条件控制。陆地高等植物叶蜡类脂物的δD与大气降水同位素的关系,及其受植被类型、蒸发和蒸腾作用等的影响是目前国际学术界关注的热点。我国学者在此方面发表了一些颇具影响的研究结果[102-105]。他们首先发现了叶蜡烷烃氢同位素能记录水汽氢同位素随高程的变化规律,进而提出了运用叶蜡烷烃氢同位素重建古高程的研究思路[102-103, 106]。水生藻类的分子标志物能记录下水体氢同位素信息,也是反映海水盐度和淡水蒸发状况的指标,但目前尚缺乏系统研究。古环境重建方面,有机氢同位素的应用已开始受到重视。
3 研究展望在未来10到20年,有机地球化学学科不仅会持续围绕着石油天然气的生成与演化这一传统内容,还将受地球学科研究趋势影响,与分支学科交叉发展。笔者认为关于过渡型有机质和成藏过程定量化重建研究将是有机地球化学研究有可能出现新突破的领域。当然,跟其他学科发展规律一样,新技术、新方法的应用也会促使学科较快的发展。因此,本节也将与有机地球化学相关的、具有良好应用前景的新仪器和新方法做一展望。
3.1 过渡型有机质过渡型有机质指的是生物有机质在向沉积有机质迁移、转化的过程中存在的一类中间态有机质(图 2)。传统的有机地球化学研究主要关注沉积有机质和生物有机质。过渡型有机质的研究还未引起人们的重视。

地球上有机质的大量形成和分布是生命出现以来生物利用CO2自我复制而不断积累的结果。其中光合作用是决定生物合成有机质的重要因素。生物体内有机化合物由4种基本类型组成:蛋白质、碳水化合物、脂类和核酸。这些化合物都含有4个最普通的元素:碳、氢、氮和氧,且在生物体内都以大分子化合物的形式存在。当然,除了这4种有机化合物外,还有一些对于维持生物机能十分重要的化合物,如维生素、单宁等。生物有机质的研究在地球生命起源、生物进化方面发挥了重要作用。通过对生物有机质的研究即是了解主体生产力的状况,也是进行沉积有机质研究的基础。
图 2显示了过渡型有机质研究的重要性。从图 2可以看出,通过对沉积有机质和生物有机质的研究,可以直接了解地球表层各圈层稳定碳的转化与循环,如,从生物有机质到沉积有机质,埋藏了多少有机碳?降解了多少有机碳?有机碳降解的速率多快?近年来,国内外在围绕碳循环的沉积有机质和生物有机质研究上取得了一些重要进展[108]。这些研究不但从另一个侧面为生命起源和生物进化的研究提供了新的事实依据,而且也为沉积环境、古气候的研究等积累了大量资料。但从生物有机质向沉积有机质迁移、转化的过程中有机质的数量及成分变化如何?各类成分变化的速率怎样?当前的有机地球化学研究并没有涉及到。要准确回答这些问题,就需要进行“过渡型有机质”的研究。
由于“过渡型有机质”是一类中间态有机质,其研究方法有别于传统生物有机质和沉积有机质。首先需要有先进的分离测定技术。同时由于目前对这些有机质的具体形成机制尚不明了,所以未来一段时间内也只能借助沉积有机质和生物有机质的研究方法对其进行表征和描述。关于“过渡型有机质”的定性与定量研究将会是未来有机地球化学学科一个新的生长点。
3.2 成藏过程的定量化重建油气生成、运移、聚集、成藏的机理和过程一直是石油地质学研究的核心,是指导勘探方向、预测成藏部位、进行油气资源评价的基础,也是油气地质—地球化学家所关注的重要内容。涉及到油气的生成、排烃与运移、聚集、后期演化等地球化学过程及其时—空演化。一般而言,对烃源岩、储集层、盖层及圈闭等油气成藏静态地质要素的认识相对容易,但对油气运聚和成藏过程等动态要素的研究则比较困难。尽管成藏过程的定量化重建面临诸多挑战,但近年来在油气成藏过程中的几个重要方面都取得长足发展,使得成藏过程的定量化重建很可能成为有机地球化学新的生长点和将来的发展方向。
成藏过程的定量化重建,需要生烃过程的定量化、排烃时间及排烃组分的定量化、充注时间的约束、聚集史及聚集过程的定量化。这些过程的定量化及其时—空匹配,构成一套完整的成藏过程重建工作,是一项地质—地球化学的综合性研究。
3.3 新的成份与同位素测定新技术先进仪器和测试技术的应用是推动有机地球化学发展的主要因素。近年来,有机化合物分析仪器的水平有了长足的发展,出现了一些分离分析能力强大的仪器分析手段,必将大大推进了有机地球化学相关研究领域的发展。
3.3.1 傅里叶变换—离子回旋共振质谱仪(FT-ICR MS)根据化合物的极性,原油和沉积岩(物)中的可溶有机质可以分为非极性的烃类组分和极性的非烃组分。烃类化合物的研究已经较为深入,常规仪器分析手段如GC、GC-MS和LC-MS等已经成为研究这一类化合物的成熟方法。与非极性的烃类化合物相比,极性化合物具有组成复杂、极性强、分子量大,难气化、易分解的特点,分离分析非常困难。电喷雾电离(ESI)—傅里叶变换离子回旋共振质谱(FT-ICR MS)是一种具有超高质量精确度和分辨率的质谱仪,可以实现石油和沉积物里可溶有机质中极性化合物复杂质谱峰的完全分离分析,并根据精确质量确定分子的元素组成。结合电喷雾(ESI)、大气压光致电离(APPI)和大气压化学电离(APCI)基质辅助激光解吸离子化(MALDI)等软电离技术,可以直接实现对原油和沉积有机质中极性化合物分子组成的分析。
3.3.2 全二维气相色谱(GC×GC)通过线性程序升温和固定相极性的改变这两者的共同作用,全二维气相色谱(GC×GC)能够实现对复杂的原油和沉积物中的可溶有机质的正交分离。具体而言,它是将分离机理不同而又互相独立的两根色谱柱以串连的方式组合成二维气相色谱。第1根色谱柱分离后的每一个馏分,经调制器聚焦后以脉冲方式进入第2根色谱柱并得到进一步的分离,通过温度和极性的改变实现气相色谱分离特性的正交化。
根据分析目的不同,GC×GC能够实现可溶有机质的族分离和目标化合物分离。根据化合物所属类型,GC×GC谱图被明显地分割成不同的区带,每一区带代表特定的族,同一族化合物在其区带内按照沸点大小不同进行分离,如烷烃、环烷烃、单环芳烃和多环芳烃等分别分布在不同的区带内,这就是GC×GC的族分离。目标化合物分离则需要进一步将感兴趣的组分与其他组分及基体进行有效分离。GC×GC因分辨率高,其定性结果比一维色谱更可靠。GC×GC也能对化合物实现定量分析,其定量分析的方法与一维色谱类似。但GC×GC作为一个全新的色谱技术,有许多难题仍需攻关,相信随着GC×GC仪器的进一步发展和完善,该技术将在复杂样品的分离和检测方面发挥举足轻重的作用。
3.3.3 放射性碳同位素(14C)测定技术随着加速质谱技术的发展,天然放射性碳(14C)作为一种重要的示踪元素及测年手段被广泛应用于考古学、生物地球化学、环境科学、海洋科学、沉积学和古气候学等研究领域,如总有机质的14C常被用于沉积有机质的测年及其沉积速率的推算。在环境科学中,随着碳质气溶胶组分分离技术的进步,对有机碳(OC)和黑碳(BC)等组分中的14C的研究获得重要进展。联合采用14C和13C技术能够为解决多种排放源的区分问题提供重要的手段,在大气碳质气溶胶源解析中具有不可替代的独特优势[109]。以往14C的测定对象多采用总有机质,但由于总有机质的组成和来源复杂,可能导致测定的14C结果难以解析,其14C年龄并不能反映沉积形成的真实年龄。生物标志物具有来源明确、示踪性强的特点,近年来,其单体分子的放射性碳同位素分析(CSRA)技术发展良好。该技术将所需的生物标志物单体分子从复杂的环境样品基质中分离并富集,再进行加速质谱仪的放射性14C测定。这种分子水平的放射性碳同位素测定技术能够揭示出总有机质同位素组成的异质性,为解释有机碳的来源、迁移和转化等提供了新型的手段,在诸多领域获得了较好的应用。未来随着单体分子分离技术的改进及加速器质谱灵敏度的提高,CSRA技术的应用会更加广泛。
3.3.4 簇同位素测定技术簇同位素方法的建立和应用是稳定同位素地球化学的一个新的突破,美国加州理工大学Eiler研究组[110-111]在簇同位素方法方面开展了一系列开创性工作。传统的同位素测定方法中每次只测定其中的一个同位素成分,如分别测定甲烷的13C成分或D成分,得到δ13C或δD的值,是为一元同位素。而簇同位素(英文称为“Clumped isotope”)的成分可以指示更精确、更方便的信息,如甲烷中13C和一个D原子同时键接在一起形成13C H3D的分子的浓度。簇同位素的浓度一般都非常小,如13C H3D的浓度比13C H4低了约三个数量级,13C H2D2的浓度比13C H3D约低六个数量级。因此簇元同位素测定需要精度极高的同位素质谱才能完成,且以往的研究多集中在碳酸岩、甲烷(CH4)和二氧化碳等组成较简单的小分子化合物。在形成碳酸盐或甲烷时,不同的温度会形成不同的簇同位素的浓度,温度越低,含有簇同位素的化合物越稳定,因而其比例也越大。因此只需要知道目标化合物本身中二元同位素占的比例,就可以确定化合物的形成温度,从而避免了需要知道生成天然气的母体有机质的同位素组成这一难题。在古海水氧位素未知的前提下,Eiler[110]的研究组最早用碳酸岩中13C-18O(Δ47) 的簇同位素的浓度来预测碳酸盐的形成温度。Quade et al.[112]和季顺川等[113]利用碳酸盐的簇同位素方法分别对全球多个地区土壤碳酸盐古温度进行了恢复重建。Suarez et al.[114]对我国黄土高原的蓝田和保德地区2~7.5 Ma的研究结果显示,碳酸盐簇同位素温度代用指标得出的古土壤平均温度都与当地现代气象站观测到的(JJSA)平均大气温度类似。也有不少学者利用甲烷的簇同位素来判断甲烷的形成温度和成因[115]。随着高精度同位素质谱技术的发展,簇同位素的应用必将越来越广泛。
致谢: 文中的部分内容据《中国地球化学学科发展史研究》第二十一章改编,对其作者表示感谢。| [1] | Rontani J F, Charrière B, Sempéré R, et al. Degradation of sterols and terrigenous organic matter in waters of the Mackenzie Shelf, Canadian Arctic[J]. Organic Geochemistry, 2014, 75: 61–73. DOI: 10.1016/j.orggeochem.2014.06.002 |
| [2] | Karlsson E S, Brüchert V, Tesi T, et al. Contrasting regimes for organic matter degradation in the East Siberian Sea and the Laptev Sea assessed through microbial incubations and molecular markers[J]. Marine Chemistry, 2015, 170: 11–22. DOI: 10.1016/j.marchem.2014.12.005 |
| [3] | Kolchugin A N, Immenhauser A, Walter B F, et al. Diagenesis of the palaeo-oil-water transition zone in a Lower Pennsylvanian carbonate reservoir:Constraints from cathodoluminescence microscopy, microthermometry, and isotope geochemistry[J]. Marine and Petroleum Geology, 2016, 72: 45–61. DOI: 10.1016/j.marpetgeo.2016.01.014 |
| [4] | Simoneit B R T, Otto A, Kusumoto N, et al. Biomarker compositions of Glyptostrobus and Metasequoia (Cupressaceae) fossils from the Eocene Buchanan Lake Formation, Axel Heiberg Island, Nunavut, Canada reflect diagenesis from terpenoids of their related extant species[J]. Review of Palaeobotany and Palynology, 2016, 235: 81–93. DOI: 10.1016/j.revpalbo.2016.07.012 |
| [5] | Alleon J, Bernard S, Le Guillou C, et al. Molecular preservation of 1. 88 Ga Gunflint organic microfossils as a function of temperature and mineralogy[J]. Nature Communications, 2016, 7: 11977. DOI: 10.1038/ncomms11977 |
| [6] | Brassell S, Eglinton G, Marlowe I, et al. Molecular stratigraphy:A new tool for climatic assessment[J]. Nature, 1986, 320(6058): 129–133. DOI: 10.1038/320129a0 |
| [7] | Brasell S C, Brereton R G, Eglinton G, et al. Palaeoclimatic signals recognized by chemometric treatment of molecular stratigraphic data[J]. Organic Geochemistry, 1986, 10(4/6): 649–660. |
| [8] | Furota S, Nakamura H, Sawada K. Long-chain alkenones and related distinctive compounds in the late Miocene and Pliocene sediments from the Gulf of Cadiz, eastern North Atlantic[J]. Organic Geochemistry, 2016, 101: 166–175. DOI: 10.1016/j.orggeochem.2016.08.014 |
| [9] | Schouten S, Hopmans E C, Schefuß E, et al. Distributional variations in marine crenarchaeotal membrane lipids:a new tool for reconstructing ancient sea water temperatures?[J]. Earth and Planetary Science Letters, 2002, 204(1/2): 265–274. |
| [10] | Ingalls A E. Signal from the subsurface[J]. Nature Geoscience, 2016, 9(8): 572–573. DOI: 10.1038/ngeo2765 |
| [11] | Ho S L, Laepple T. Flat meridional temperature gradient in the early Eocene in the subsurface rather than surface ocean[J]. Nature Geoscience, 2016, 9(8): 606–610. DOI: 10.1038/ngeo2763 |
| [12] | Rampen S W, Willmott V, Kim J H, et al. Long chain 1, 13-and 1, 15-diols as a potential proxy for palaeotemperature reconstruction[J]. Geochimica et Cosmochimica Acta, 2012, 84: 204–216. DOI: 10.1016/j.gca.2012.01.024 |
| [13] | Rampen S W, Willmott V, Kim J H, et al. Evaluation of long chain 1, 14-alkyl diols in marine sediments as indicators for upwelling and temperature[J]. Organic Geochemistry, 2014, 76: 39–47. DOI: 10.1016/j.orggeochem.2014.07.012 |
| [14] | De Bar M W, Dorhout D J C, Hopmans E C, et al. Constraints on the application of long chain diol proxies in the Iberian Atlantic margin[J]. Organic Geochemistry, 2016, 101: 184–195. DOI: 10.1016/j.orggeochem.2016.09.005 |
| [15] | Dos Santos R A L, Spooner M I, Barrows T T, et al. Comparison of organic (U37K', TEX86H, LDI) and faunal proxies (foraminiferal assemblages) for reconstruction of late Quaternary sea surface temperature variability from offshore southeastern Australia[J]. Paleoceanography, 2013, 28(3): 377–387. DOI: 10.1002/palo.20035 |
| [16] | Rodrigo-Gámiz M, Martínez-Ruiz F, Rampen S W, et al. Sea surface temperature variations in the western Mediterranean Sea over the last 20 kyr:A dual-organic proxy (U37K' and LDI) approach[J]. Paleoceanography, 2014, 29(2): 87–98. DOI: 10.1002/2013PA002466 |
| [17] | Becker K W, Lipp J S, Versteegh J G M, et al. Rapid and simultaneous analysis of three molecular sea surface temperature proxies and application to sediments from the Sea of Marmara[J]. Organic Geochemistry, 2015, 85: 42–53. DOI: 10.1016/j.orggeochem.2015.04.008 |
| [18] | Sigman D M, Boyle E A. Glacial/interglacial variations in atmospheric carbon dioxide[J]. Nature, 2000, 407(6806): 859–869. DOI: 10.1038/35038000 |
| [19] | Falkowski P, Scholes R J, Boyle E, et al. The global carbon cycle:A test of our knowledge of earth as a system[J]. Science, 2000, 290(5490): 291–296. DOI: 10.1126/science.290.5490.291 |
| [20] | Jaccard S L, Hayes C T, Martínez-García A, et al. Two modes of change in Southern Ocean productivity over the past million years[J]. Science, 2013, 339(6126): 1419–1423. DOI: 10.1126/science.1227545 |
| [21] | Kawamura K, Ishimura Y, Yamazaki K. Four years' observations of terrestrial lipid class compounds in marine aerosols from the western North Pacific[J]. Global Biogeochemical Cycles, 2003, 17(1): 3–1. DOI: 10.1029/2001GB001810 |
| [22] | Burdige D J. Burial of terrestrial organic matter in marine sediments:a re-assessment[J]. Global Biogeochemical Cycles, 2005, 19(4): GB4011. |
| [23] | Zhang Y L, Kaiser K, Li L, et al. Sources, distributions, and early diagenesis of sedimentary organic matter in the Pearl River region of the South China Sea[J]. Marine Chemistry, 2014, 158: 39–48. DOI: 10.1016/j.marchem.2013.11.003 |
| [24] | Schubert C J, Villanueva J, Calvert S E, et al. Stable phytoplankton community structure in the Arabian Sea over the past 200, 000 years[J]. Nature, 1998, 394(6693): 563–566. DOI: 10.1038/29047 |
| [25] | McMahon K W, McCarthy M D, Sherwood O A, et al. Millennial-scale plankton regime shifts in the subtropical North Pacific Ocean[J]. Science, 2015, 354(6267): 1530–1533. |
| [26] | Lourenço R A, De Mahiques M M, Wainer I E K C, et al. Organic biomarker records spanning the last 34, 800 years from the southeastern Brazilian upper slope:links between sea surface temperature, displacement of the Brazil Current, and marine productivity[J]. Geo-Marine Letters, 2016, 36(5): 361–369. DOI: 10.1007/s00367-016-0453-7 |
| [27] | Weijers J W H, Schouten S, Van Den Donker J C, et al. Environmental controls on bacterial tetraether membrane lipid distribution in soils[J]. Geochimica et Cosmochimica Acta, 2007, 71(3): 703–713. DOI: 10.1016/j.gca.2006.10.003 |
| [28] | Loomis S E, Russell J M, Ladd B, et al. Calibration and application of the branched GDGT temperature proxy on East African lake sediments[J]. Earth and Planetary Science Letters, 2012, 357-358: 277–288. DOI: 10.1016/j.epsl.2012.09.031 |
| [29] | Schouten S, Hopmans E C, Damsté J S S. The organic geochemistry of glycerol dialkyl glycerol tetraether lipids:a review[J]. Organic Geochemistry, 2013, 54: 19–61. DOI: 10.1016/j.orggeochem.2012.09.006 |
| [30] | Crump B C, Amaral-Zettler L, Kling G W. Microbial diversity in arctic freshwaters is structured by inoculation of microbes from soils[J]. The ISME Journal, 2012, 6(9): 1629–1639. DOI: 10.1038/ismej.2012.9 |
| [31] | Theroux S, D'Andrea W J, Toney J, et al. Phylogenetic diversity and evolutionary relatedness of alkenone-producing haptophyte algae in lakes:implications for continental paleotemperature reconstructions[J]. Earth and Planetary Science Letters, 2010, 300(3/4): 311–320. |
| [32] | D'Andrea W J, Theroux S, Bradley R S, et al. Does phylogeny control U37K-temperature sensitivity? Implications for lacustrine alkenone paleothermometry[J]. Geochimica et Cosmochimica Acta, 2016, 175: 168–180. DOI: 10.1016/j.gca.2015.10.031 |
| [33] | Simon M, López-García P, Moreira D, et al. New haptophyte lineages and multiple independent colonizations of freshwater ecosystems[J]. Environmental Microbiology Reports, 2013, 5(2): 322–332. DOI: 10.1111/emi4.2013.5.issue-2 |
| [34] | McColl J L, Couto J, Bendle J A, et al. 18S rDNA analysis of alkenone-producing haptophyte(s) preserved in surface sediments of Lake Toyoni, Japan[C]//American Geophysical Union Fall Meeting 2013. Washington:AGU, 2013. http://adsabs.harvard.edu/abs/2013agufmpp43b2084m |
| [35] | Longo W M, Theroux S, Giblin A E, et al. Temperature calibration and phylogenetically distinct distributions for freshwater alkenones:Evidence from northern Alaskan lakes[J]. Geochimica et Cosmochimica Acta, 2016, 180: 177–196. DOI: 10.1016/j.gca.2016.02.019 |
| [36] | Toney J L, Huang Y S, Fritz S C, et al. Climatic and environmental controls on the occurrence and distributions of long chain alkenones in lakes of the interior United States[J]. Geochimica et Cosmochimica Acta, 2010, 74(5): 1563–1578. DOI: 10.1016/j.gca.2009.11.021 |
| [37] | Zheng Y S, Huang Y S, Andersen R A, et al. Excluding the di-unsaturated alkenone in the U37K index strengthens temperature correlation for the common lacustrine and brackish-water haptophytes[J]. Geochimica et Cosmochimica Acta, 2016, 175: 36–46. DOI: 10.1016/j.gca.2015.11.024 |
| [38] | Bird B W, Polisar P J, Lei Y B, et al. A Tibetan lake sediment record of Holocene Indian summer monsoon variability[J]. Earth and Planetary Science Letters, 2014, 399: 92–102. DOI: 10.1016/j.epsl.2014.05.017 |
| [39] | Günther F, Witt R, Schouten S, et al. Quaternary ecological responses and impacts of the Indian Ocean Summer Monsoon at Nam Co, Southern Tibetan Plateau[J]. Quaternary Science Reviews, 2015, 112: 66–77. DOI: 10.1016/j.quascirev.2015.01.023 |
| [40] | Sessions A L. Factors controlling the deuterium contents of sedimentary hydrocarbons[J]. Organic Geochemistry, 2016, 96: 43–64. DOI: 10.1016/j.orggeochem.2016.02.012 |
| [41] | O'Leary M H. Carbon isotope fractionation in plants[J]. Phytochemistry, 1981, 20(4): 553–567. DOI: 10.1016/0031-9422(81)85134-5 |
| [42] | Garcin Y, Schwab V F, Gleixner G, et al. Hydrogen isotope ratios of lacustrine sedimentary n-alkanes as proxies of tropical African hydrology:insights from a calibration transect across Cameroon[J]. Geochimica et Cosmochimica Acta, 2012, 79: 106–126. DOI: 10.1016/j.gca.2011.11.039 |
| [43] | Yang S L, Ding Z L, Li Y Y, et al. Warming-induced northwestward migration of the East Asian monsoon rain belt from the Last Glacial Maximum to the mid-Holocene[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(43): 13178–13183. DOI: 10.1073/pnas.1504688112 |
| [44] | Garcin Y, Schefuß E, Schwab V F, et al. Reconstructing C3 and C4 vegetation cover using n-alkane carbon isotope ratios in recent lake sediments from Cameroon, Western Central Africa[J]. Geochimica et Cosmochimica Acta, 2014, 142: 482–500. DOI: 10.1016/j.gca.2014.07.004 |
| [45] | Sage R F, Sage T L, Kocacinar F. Photorespiration and the evolution of C4 photosynthesis[J]. Annual Review of Plant Biology, 2012, 63(1): 19–47. DOI: 10.1146/annurev-arplant-042811-105511 |
| [46] | Vicentini A, Barber J C, Aliscioni S S, et al. The age of the grasses and clusters of origins of C4 photosynthesis[J]. Global Change Biology, 2008, 14(12): 2963–2977. DOI: 10.1111/gcb.2008.14.issue-12 |
| [47] | Hoetzel S, Dupont L, Schefuß E, et al. The role of fire in Miocene to Pliocene C4 grassland and ecosystem evolution[J]. Nature Geoscience, 2013, 6(12): 1027–1030. DOI: 10.1038/ngeo1984 |
| [48] | Nelson D M, Urban M A, Kershaw A P, et al. Late-Quaternary variation in C3 and C4 grass abundance in southeastern Australia as inferred from δ13C analysis:Assessing the roles of climate, pCO2, and fire[J]. Quaternary Science Reviews, 2016, 139: 67–76. DOI: 10.1016/j.quascirev.2016.03.006 |
| [49] | Castañeda I S, Schouten S. A review of molecular organic proxies for examining modern and ancient lacustrine environments[J]. Quaternary Science Reviews, 2011, 30(21/22): 2851–2891. |
| [50] | Diefendorf A F, Mueller K E, Wing S L, et al. Global patterns in leaf 13C discrimination and implications for studies of past and future climate[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(13): 5738–5743. DOI: 10.1073/pnas.0910513107 |
| [51] | Diefendorf A F, Leslie A B, Wing S L. Leaf wax composition and carbon isotopes vary among major conifer groups[J]. Geochimica et Cosmochimica Acta, 2015, 170: 145–156. DOI: 10.1016/j.gca.2015.08.018 |
| [52] | Eley Y, Dawson L, Pedentchouk N. Investigating the carbon isotope composition and leaf wax n-alkane concentration of C3 and C4 plants in Stiffkey saltmarsh, Norfolk, UK[J]. Organic Geochemistry, 2016, 96: 28–42. DOI: 10.1016/j.orggeochem.2016.03.005 |
| [53] | Van Bentum E C, Hetzel A, Brumsack H J, et al. Reconstruction of water column anoxia in the equatorial Atlantic during the Cenomanian-Turonian oceanic anoxic event using biomarker and trace metal proxies[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2009, 280(3/4): 489–498. |
| [54] | Hayes J M, Freeman K H, Brian N, et al. Compound-specific isotopic analyses:A novel tool for reconstruction of ancient biogeochemical processes[J]. Organic Geochemistry, 1990, 16(4/5/6): 1115–1128. |
| [55] | Wakeham S G, Amann R, Freeman K H, et al. Microbial ecology of the stratified water column of the Black Sea as revealed by a comprehensive biomarker study[J]. Organic Geochemistry, 2007, 38(12): 2070–2097. DOI: 10.1016/j.orggeochem.2007.08.003 |
| [56] | Lu H, Sheng G Y, Peng P A, et al. Identification of C24 and C25 lanostanes in Tertiary sulfur rich crude oils from the Jinxian Sag, Bohai Bay Basin, Northern China[J]. Organic Geochemistry, 2011, 42(2): 146–155. DOI: 10.1016/j.orggeochem.2010.11.009 |
| [57] | Zhang C M, Zhang Y Q, Cai C F. Aromatic isoprenoids from the 25-65 Ma saline lacustrine formations in the western Qaidam Basin, NW China[J]. Organic Geochemistry, 2011, 42(7): 851–855. DOI: 10.1016/j.orggeochem.2011.04.010 |
| [58] | Lu H, Shen C C, Zhang Z R, et al. 2, 3, 6-/2, 3, 4-Aryl isoprenoids in Paleocene crude oils from Chinese Jianghan Basin:Constrained by water column stratification[J]. Energy & Fuels, 2015, 29(8): 4690–4700. |
| [59] | Tian H, Cheng P, Zhou Q, et al. A complete series of C31-C34 25-norbenzohopanes in the Devonian and Jurassic bitumen sands, NW Sichuan Basin[J]. Organic Geochemistry, 2012, 45: 1–6. DOI: 10.1016/j.orggeochem.2012.01.003 |
| [60] | 李敏, 田辉, 肖贤明, 等. 塔河油田稠油中25-降苯并藿烷的检出及其意义[J]. 地球化学, 2014, 43 (5): 453–459. [ Li Min, Tian Hui, Xiao Xianming, et al. Identification of 25-norbenzohopanes in Tahe oilfield and its geological significance[J]. Geochimica, 2014, 43(5): 453–459. ] |
| [61] | 李美俊, 师生宝, 王铁冠, 等. 石油和沉积有机质中C3-、C4-烷基取代二苯并噻吩的鉴定[J]. 地球化学, 2014, 43 (2): 157–165. [ Liu Meijun, Shi Shengbao, Wang Tieguan, et al. The identification of C3-and C4-alkylated dibenzothiophenes in petroleum and sedimentary organic matter[J]. Geochimica, 2014, 43(2): 157–165. ] |
| [62] | Li M J, Wang T G, Shi S B, et al. Benzo[b]naphthothiophenes and alkyl dibenzothiophenes:Molecular tracers for oil migration distances[[J]. Marine and Petroleum Geology, 2014, 57: 403–417. DOI: 10.1016/j.marpetgeo.2014.06.012 |
| [63] | 傅家谟, 刘德汉, 盛国英. 煤成烃地球化学[M]. 北京: 科学出版社, 1990: 1-372. [ Fu Jiamo, Liu Dehan, Sheng Guoying. Coal-generated hydrocarbon geochemistry[M]. Beijing: Science Press, 1990: 1-372. ] |
| [64] | 戴金星, 倪云燕, 黄士鹏, 等. 煤成气研究对中国天然气工业发展的重要意义[J]. 天然气地球科学, 2014, 25 (1): 1–22. [ Dai Jingxing, Ni Yunyan, Huang Shipeng, et al. Significant function of coal-derived gas study for natural gas industry development in China[J]. Natural Gas Geoscience, 2014, 25(1): 1–22. ] |
| [65] | 戴金星. 中国煤成气研究30年来勘探的重大进展[J]. 石油勘探与开发, 2009, 36 (2): 264–279. [ Dai Jingxing. Major developments of coal-formed gas exploration in the last 30 years in China[J]. Petroleum Exploration and Development, 2009, 36(2): 264–279. ] |
| [66] | 姜在兴, 肖尚斌. 渤海湾盆地深部油气资源[J]. 勘探家, 1998, 3 (1): 16–19. [ Jiang Zaixing, Xiao Shangbin. Deep oil-gas resource in Bohaiwan Basin[J]. Petroleum Explorationist, 1998, 3(1): 16–19. ] |
| [67] | 李小地. 中国深部油气藏的形成与分布初探[J]. 石油勘探与开发, 1994, 21 (1): 34–39, 125. [ Li Xiaodi. Formation and distribution of deep oil-gas reservoir in China[J]. Petroleum Exploration and Development, 1994, 21(1): 34–39, 125. ] |
| [68] | 邹才能, 杜金虎, 徐春春, 等. 四川盆地震旦系-寒武系特大型气田形成分布、资源潜力及勘探发现[J]. 石油勘探与开发, 2014, 41 (1): 278–293. [ Zou Caineng, Du Jinhu, Xu Chunchun, et al. Formation, distribution, resource potential and discovery of the Sinian-Cambrian giant gas field, Sichuan Basin, SW China[J]. Petroleum Exploration and Development, 2014, 41(1): 278–293. ] |
| [69] | 刘文汇, 王杰, 腾格尔, 等. 中国海相层系多元生烃及其示踪技术[J]. 石油学报, 2012, 33 (Suppl.1): 115–125. [ Liu Wenhui, Wang Jie, Tenger, et al. Multiple hydrocarbon generation of marine strata and its tracer technique in China[J]. Acta Petrolei Sinica, 2012, 33(Suppl.1): 115–125. ] |
| [70] | 庞雄奇, 周新源, 姜振学, 等. 叠合盆地油气藏形成、演化与预测评价[J]. 地质学报, 2012, 86 (1): 1–103. [ Pang Xiongqi, Zhou Xinyuan, Jiang Zhenxue, et al. Hydrocarbon reservoirs formation, evolution, prediction and evaluation in the superimposed basins[J]. Acta Geologica Sinica, 2012, 86(1): 1–103. ] |
| [71] | 王招明. 塔里木盆地库车坳陷克拉苏盐下深层大气田形成机制与富集规律[J]. 天然气地球科学, 2014, 25 (2): 153–166. [ Wang Zhaoming. Formation mechanism and enrichment regularities of Kelasu subsalt deep large gas field in Kuqu depression, Tarim Basin[J]. Natural Gas Geoscience, 2014, 25(2): 153–166. ] |
| [72] | 张水昌, 张斌, 杨海军, 等. 塔里木盆地喜马拉雅晚期油气藏调整与改造[J]. 石油勘探与开发, 2012, 39 (6): 668–680. [ Zhang Shuicang, Zhang Bin, Yang Haijun, et al. Adjustment and alteration of hydrocarbon reservoirs during the late Himalayan Period, Tarim Basin, NW China[J]. Petroleum Exploration and Development, 2012, 39(6): 668–680. ] |
| [73] | 赵文智, 朱光有, 苏劲, 等. 中国海相油气多期充注与成藏聚集模式研究:以塔里木盆地轮古东地区为例[J]. 岩石学报, 2012, 28 (3): 709–721. [ Zhao Wenzhi, Zhu Guangyou, Su Jin, et al. Study on the multi-stage charging and accumulation model of chinese marine petroleum:Example from eastern Lungu area in the Tarim Basin[J]. Acta Petrologica Sinica, 2012, 28(3): 709–721. ] |
| [74] | Sun Q, Chu G Q, Liu G X, et al. Calibration of alkenone unsaturation index with growth temperature for a lacustrine species, Chrysotila lamellosa (Haptophyceae)[J]. Organic Geochemistry, 2007, 38(8): 1226–1234. DOI: 10.1016/j.orggeochem.2007.04.007 |
| [75] | Song M, Zhou A F, He Y X, et al. Environmental controls on long-chain alkenone occurrence and compositional patterns in lacustrine sediments, northwestern China[J]. Organic Geochemistry, 2016, 91: 43–53. DOI: 10.1016/j.orggeochem.2015.10.009 |
| [76] | Dang X Y, Yang H, Naafs B D A, et al. Evidence of moisture control on the methylation of branched glycerol dialkyl glycerol tetraethers in semi-arid and arid soils[J]. Geochimica et Cosmochimica Acta, 2016, 189: 24–36. DOI: 10.1016/j.gca.2016.06.004 |
| [77] | Hu J F, Zhou H D, Peng P A, et al. Seasonal variability in concentrations and fluxes of glycerol dialkyl glycerol tetraethers in Huguangyan Maar Lake, SE China:Implications for the applicability of the MBT-CBT paleotemperature proxy in lacustrine settings[J]. Chemical Geology, 2016, 420: 200–212. DOI: 10.1016/j.chemgeo.2015.11.008 |
| [78] | Wang J X, Wei Y L, Wang P, et al. Unusually low TEX86 values in the transitional zone between Pearl River estuary and coastal South China Sea:Impact of changing archaeal community composition[J]. Chemical Geology, 2015, 402: 18–29. DOI: 10.1016/j.chemgeo.2015.03.002 |
| [79] | Wu W C, Ruan J P, Ding S, et al. Source and distribution of glycerol dialkyl glycerol tetraethers along lower Yellow River-estuary-coast transect[J]. Marine Chemistry, 2014, 158: 17–26. DOI: 10.1016/j.marchem.2013.11.006 |
| [80] | Yang H, Pancost R D, Dang X Y, et al. Correlations between microbial tetraether lipids and environmental variables in Chinese soils:Optimizing the paleo-reconstructions in semi-arid and arid regions[J]. Geochimica et Cosmochimica Acta, 2014, 126: 49–69. DOI: 10.1016/j.gca.2013.10.041 |
| [81] | Zhou H D, Hu J F, Spiro B, et al. Glycerol dialkyl glycerol tetraethers in surficial coastal and open marine sediments around China:Indicators of sea surface temperature and effects of their sources[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 395: 114–121. DOI: 10.1016/j.palaeo.2013.12.006 |
| [82] | Chu G Q, Sun Q, Wang X H, et al. Seasonal temperature variability during the past 1600 years recorded in historical documents and varved lake sediment profiles from northeastern China[J]. Holocene, 2012, 22(7): 785–792. DOI: 10.1177/0959683611430413 |
| [83] | He Y X, Zhao C, Wang Z, et al. Late Holocene coupled moisture and temperature changes on the northern Tibetan Plateau[J]. Quaternary Science Reviews, 2013, 80: 47–57. DOI: 10.1016/j.quascirev.2013.08.017 |
| [84] | Wang Z, Liu Z H, Zhang F, et al. A new approach for reconstructing Holocene temperatures froma multi-species long chain alkenone record from Lake Qinghai on the northeastern Tibetan Plateau[J]. Organic Geochemistry, 2015, 88: 50–58. DOI: 10.1016/j.orggeochem.2015.08.006 |
| [85] | Ling Y, Sun Q, Zheng M P, et al. Alkenone-based temperature and climate reconstruction during the last deglaciation at Lake Dangxiong Co, southwestern Tibetan Plateau[J]. Quaternary International, 2017, 443: 58–69. DOI: 10.1016/j.quaint.2016.07.036 |
| [86] | Li D W, Zhao M X, Chen M T. East Asian winter monsoon controlling phytoplankton productivity and community structure changes in the southeastern South China Sea over the last 185 kyr[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 414: 233–242. DOI: 10.1016/j.palaeo.2014.09.003 |
| [87] | Kong D M, Zong Y Q, Jia G D, et al. The development of late Holocene coastal cooling in the northern South China Sea[J]. Quaternary International, 2014, 349: 300–307. DOI: 10.1016/j.quaint.2013.08.055 |
| [88] | Tao S Q, Xing L, Luo X F, et al. Alkenone distribution in surface sediments of the southern Yellow Sea and implications for the U37K' thermometer[J]. Geo-Marine Letters, 2012, 32(1): 61–71. DOI: 10.1007/s00367-011-0251-1 |
| [89] | Zhang J, Bai Y, Xu S D, et al. Alkenone and tetraether lipids reflect different seasonal seawater temperatures in the coastal northern South China Sea[J]. Organic Geochemistry, 2013, 58: 115–120. DOI: 10.1016/j.orggeochem.2013.02.012 |
| [90] | Hu J F, Zhou H D, Peng P A, et al. Reconstruction of a paleotemperature record from 0. 3-3. 7 ka for subtropical South China using lacustrine branched GDGTs from Huguangyan Maar[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2015, 435: 167–176. DOI: 10.1016/j.palaeo.2015.06.014 |
| [91] | Li D W, Zhao M X, Tian J, et al. Comparison and implication of TEX86 and U37K' temperature records over the last 356 kyr of ODP Site 1147 from the northern South China Sea[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 376: 213–223. DOI: 10.1016/j.palaeo.2013.02.031 |
| [92] | Lu H X, Liu W G, Wang H Y, et al. Variation in 6-methyl branched glycerol dialkyl glycerol tetraethers in Lantian loess-paleosol sequence and effect on paleotemperature reconstruction[J]. Organic Geochemistry, 2016, 100: 10–17. DOI: 10.1016/j.orggeochem.2016.07.006 |
| [93] | Wang H Y, Dong H L, Zhang C L, et al. A 12-kyr record of microbial branched and isoprenoid tetraether index in Lake Qinghai, northeastern Qinghai-Tibet Plateau:Implications for paleoclimate reconstruction[J]. Science China Earth Sciences, 2016, 59(5): 951–960. DOI: 10.1007/s11430-015-5213-4 |
| [94] | Zheng Y H, Li Q Y, Wang Z Z, et al. Peatland GDGT records of Holocene climatic and biogeochemical responses to the Asian Monsoon[J]. Organic Geochemistry, 2015, 87: 86–95. DOI: 10.1016/j.orggeochem.2015.07.012 |
| [95] | Li Y Y, Yang S L, Wang X, et al. Leaf wax n-alkane distributions in Chinese loess since the Last Glacial Maximum and implications for paleoclimate[J]. Quaternary International, 2016, 399: 190–197. DOI: 10.1016/j.quaint.2015.04.029 |
| [96] | Jia G D, Li Z Y, Peng P A, et al. Aeolian n-alkane isotopic evidence from North Pacific for a Late Miocene decline of C4 plant in the arid Asian interior[J]. Earth and Planetary Science Letters, 2012, 321-322: 32–40. DOI: 10.1016/j.epsl.2011.12.044 |
| [97] | Zhou B, Shen C D, Sun W D, et al. Late Pliocene-Pleistocene expansion of C4 vegetation in semiarid East Asia linked to increased burning[J]. Geology, 2014, 42(12): 1067–1070. DOI: 10.1130/G36110.1 |
| [98] | He J, Zhao M X, Wang P X, et al. Changes in phytoplankton productivity and community structure in the northern South China Sea during the past 260 ka[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 392: 312–323. DOI: 10.1016/j.palaeo.2013.09.010 |
| [99] | Li L, Li Q Y, He J, et al. Biomarker-derived phytoplankton community for summer monsoon reconstruction in the western South China Sea over the past 450 ka[J]. Deep-Sea Research Ⅱ:Topical Studies in Oceanography, 2015, 122: 118–130. DOI: 10.1016/j.dsr2.2015.11.006 |
| [100] | Luo G M, Wang Y B, Grice K, et al. Microbial-algal community changes during the latest Permian ecological crisis:Evidence from lipid biomarkers at Cili, South China[J]. Global and Planetary Change, 2013, 105: 36–51. DOI: 10.1016/j.gloplacha.2012.11.015 |
| [101] | Xie S C, Pancost R D, Huang X Y, et al. Molecular and isotopic evidence for episodic environmental change across the Permo/Triassic boundary at Meishan in South China[J]. Global and Planetary Change, 2007, 55(1/3): 56–65. |
| [102] | Bai Y, Fang X M, Jia G D, et al. Different altitude effect of leaf wax n-alkane δD values in surface soils along two vapor transport pathways, southeastern Tibetan Plateau[J]. Geochimica et Cosmochimica Acta, 2015, 170: 94–107. DOI: 10.1016/j.gca.2015.08.016 |
| [103] | Jia G D, Bai Y, Ma Y J, et al. Paleoelevation of Tibetan Lunpola basin in the Oligocene-Miocene transition estimated from leaf wax lipid dual isotopes[J]. Global and Planetary Change, 2015, 126: 14–22. DOI: 10.1016/j.gloplacha.2014.12.007 |
| [104] | Rao Z G, Jia G D, Qiang M R, et al. Assessment of the difference between mid-and long chain compound specific δDn-alkanes values in lacustrine sediments as a paleoclimatic indicator[J]. Organic Geochemistry, 2014, 76: 104–117. DOI: 10.1016/j.orggeochem.2014.07.015 |
| [105] | Rao Z G, Jia G D, Liu Y X, et al. Asynchronous evolution of the isotopic composition and amount of precipitation in north China during the Holocene revealed by a record of compound-specific carbon and hydrogen isotopes of long-chain n-alkanes from an alpine lake[J]. Earth and Planetary Science Letters, 2016, 446: 68–76. DOI: 10.1016/j.epsl.2016.04.027 |
| [106] | Luo P, Peng P A, Gleixner G, et al. Empirical relationship between leaf wax n-alkane dD and altitude in the Wuyi, Shennongjia and Tianshan Mountains China:implications for Paleoaltimetry[J]. Earth and Planetary Science Letters, 2011, 301(1/2): 285–296. |
| [107] | 焦念志. 海洋固碳与储碳——并论微型生物在其中的重要作用[J]. 中国科学(D辑):地球科学, 2012, 42 (10): 1473–1486. [ Jiao Nianzhi. Carbon fixation and sequestration in the ocean, with special reference to the microbial carbon pump[J]. Science China(Seri. D):Earth Sciences, 2012, 42(10): 1473–1486. ] |
| [108] | Pan Y D, Birdsey R A, Fang J Y, et al. A large and persistent carbon sink in the world's forests[J]. Science, 2011, 333(6045): 988–993. DOI: 10.1126/science.1201609 |
| [109] | 张世春, 王毅勇, 童全松. 碳同位素技术在碳质气溶胶源解析中应用的研究进展[J]. 地球科学进展, 2013, 28 (1): 62–70. [ Zhang Shichun, Wang Yiyong, Tong Quansong. The use of carbon isotope analysis in source apportionment of carbonaceous aerosols:A Review[J]. Advances Earth Science, 2013, 28(1): 62–70. DOI: 10.11867/j.issn.1001-8166.2013.01.0062 ] |
| [110] | Eiler J M. "Clumped-isotope" geochemistry-The study of naturally-occurring, multiply-substituted isotopologues[J]. Earth and Planetary Science Letters, 2007, 262(3/4): 309–327. |
| [111] | Ghosh P, Adkins J, Affek H, et al. 13C-18O bonds in carbonate minerals:A new kind of paleothermometer[J]. Geochimica et Cosmochimica Acta, 2006, 70(6): 1439–1456. DOI: 10.1016/j.gca.2005.11.014 |
| [112] | Quade J, Eiler J, Daëron M, et al. The clumped isotope geothermometer in soil and paleosol carbonate[J]. Geochimica et Cosmochimica Acta, 2013, 105: 92–107. DOI: 10.1016/j.gca.2012.11.031 |
| [113] | 季顺川, 彭廷江, 聂军胜, 等. 黄土高原微生物膜类脂物和碳酸盐二元同位素重建古温度的研究进展[J]. 海洋地质与第四纪地质, 2013, 33 (3): 151–158. [ Ji Shunchuan, Peng Tingjiang, Nie Junsheng, et al. Quantitative paleotempererature reconstruction of the Chinese loess plateau:A review[J]. Marine Geology & Quaternary Geology, 2013, 33(3): 151–158. ] |
| [114] | Suarez M B, Passey B H, Kaakinen A. Paleosol carbonate multiple isotopologue signature of active East Asian summer monsoons during the late Miocene and Pliocene[J]. Geology, 2011, 39(12): 1151–1154. DOI: 10.1130/G32350.1 |
| [115] | Stolper D A, Lawson M, Davis C L, et al. Formation temperatures of thermogenic and biogenic methane[J]. Science, 2014, 344(6191): 1500–1503. DOI: 10.1126/science.1254509 |