



动脉粥样硬化(atherosclerosis,AS)是严重危害人类健康的一种血管慢性炎症性疾病[1, 2, 3, 4]。基本病变过程是动脉内膜脂质沉积,内膜灶状纤维化,粥样斑块形成,致血管壁变厚、管腔狭窄,并引起一系列继发性病变,如引起冠心病、脑血管病和血栓性疾病等。到目前为止,AS的发病机制尚未完全清楚。普遍认为脂质代谢紊乱引起的炎症反应与AS的发生发展有密切关系[3, 5, 6]。炎症反应中,黏附分子如选择素、选择素配体、整合素等介导的单核-内皮细胞间的黏附在粥样斑块的发生发展中起了极其重要的作用[1],是AS的早期特征和关键环节。这些环节的调控与多种因素有关,蛋白质翻译后的调节是否参与其中还鲜为人知。我们团队以往的研究显示,蛋白质的糖基化对单核细胞-内皮之间相互作用的调控发挥着重要的作用[7, 8]。蛋白糖基化是由细胞中催化特殊氨基酸残基联接聚糖基团的糖基转移酶催化形成。其中β-半乳糖α2,6-唾液酸转移酶1(ST6Gal-Ⅰ)是高尔基体上一个Ⅱ型跨膜蛋白,可催化唾液酸以α-2,6糖苷键的形式连接到细胞膜表面功能蛋白的N-乙酰半乳糖胺上形成唾液酸化的糖蛋白,从而发挥生物学功能[9]。有研究表明,白细胞在PMA刺激分化成巨噬细胞过程中,ST6Gal-Ⅰ表达下调,膜上β1整合素的α2,6-唾液酸化明显降低,从而增加了细胞的α4β1整合素与VCAM-1的黏附,说明炎症刺激下ST6Gal-Ⅰ确实参与细胞黏附调控[10]。既然糖基转移酶介导的糖基化修饰影响了细胞的生物学功能,那么本文中糖基转移酶对AS过程中斑块形成是否存在调控关系就成为我们关注的核心问题。这对揭示AS的发生、发展具有重要意义。
目前,瑞舒伐他汀是有效防治AS的药物。他汀类药物主要通过减少内源性胆固醇的合成,有效降低LDL-C、升高HLD-L,还可以通过上调内皮eNOS的表达发挥抗炎作用[11]。但是瑞舒伐他汀发挥抗AS机制的深度认识尚不全面。基于AS是在各种炎症作用下发生的一系列病理改变,且炎症与ST6Gal-Ⅰ的表达密切相关。探讨药物抗AS作用与ST6Gal-Ⅰ的表达关系,对于发现新靶点具有重要意义。已有文献报道,AS病变与ST3Gal-Ⅳ密切相关[12];在炎症发生时血清中的α2,6-、α2,3-及α2,8-唾液酸化糖蛋白均发生差异表达[13]。而瑞舒伐他汀对AS的防治是否与唾液酸化糖蛋白表达有关尚未见报道。因此,本研究拟以ApoE-/-小鼠为实验对象,高脂饲料喂养建立AS动物模型,分析瑞舒伐他汀药物对AS斑块的作用,并观察给药前后ST6Gal-Ⅰ在ApoE-/-小鼠血管AS形成区域的表达。以期判断瑞舒伐他汀治疗AS机制是否与功能蛋白糖基化有关。
1 材料与方法 1.1 动物和试剂健康♂载脂蛋白E基因敲除小鼠(ApoE knocked-out mice ,ApoE-/-),6周龄,体质量18~25 g,购置于重庆医科大学实验动物中心。瑞舒伐他汀(博腾精细化工股份有限公司,重庆);油红O(Sigma,USA);苏木精、伊红、兔超敏二步法免疫组化检测试剂盒、DAB染色试剂盒(中杉金桥,北京);兔源多克隆ST6Gal-Ⅰ抗体(Sigma,USA);荧光正置显微镜(Olympus,日本);血糖试纸(罗氏,德国);高脂高胆固醇饲料(含20%脂肪,0.125%胆固醇)由北京科澳协力饲料有限公司制备。
1.2 动物模型的构建及分组36只ApoE-/-小鼠于SPF级分笼喂养,应用随机表原则,将其分为3组,每组12只:① 基线模型组(baseline):髙脂高胆固醇饲料喂养16周处死;② 空白对照组(control):髙脂高胆固醇饲料喂养23周处死;③ 瑞舒伐他汀组(rosuvastatin):高脂高胆固醇饲料喂养23周,并在后7周同时对小鼠给予瑞舒伐他汀灌胃处理,瑞舒伐他汀以PBS缓冲液配置,给药剂量2.4 mg·kg-1·d-1[14]。处死前1 d晚上禁食不禁水。
1.3 血样的采集及生化指标的测定ApoE-/-小鼠经戊巴比妥麻醉后行眼眶采血,分离血清,测定血糖、血脂四项,即低密度脂蛋白(LDL-C)、高密度脂蛋白(HDL-C)、总胆固醇(TC)、甘油三酯(TG)的水平,于重庆医科大学附属大学城医院检验科测得。
1.4 标本的采集及主动脉标本的制备ApoE-/-小鼠麻醉采血后处死,剖开胸腹腔用生理盐水冲洗干净,分离心、肝、脾、肺、肾、小肠及心脏等重要脏器,取出心脏和主动脉,从主动脉根部开始取材,剪下1~1.5 cm连接心脏的血管放入4%的多聚甲醛中固定,经洗涤、脱水、透明,进行常规石蜡包埋,从主动脉根部0.2 cm起间断均匀切面,切片厚度为5 μm,用防脱玻片黏附切片。
1.5 油红O染色取主动脉弓(1~2 mm)的心脏,剔除动脉弓处多余脂肪,充分暴露主动脉,横切左右心房,是切面垂直主动脉根部,用吸水纸将组织块的固定液充分吸干,OCT包埋剂滴到样品托盘上,是主动脉垂直朝上将心房组织完全植入到包埋剂中。调整组织使主动脉垂直向上,将样品连同托盘放到速冻台上迅速冰冻30 min。冰冻切片机切片,温度-20℃左右,切片厚度8 μm,使用防脱玻片不同位置贴片,并在普通显微镜10倍视野下,观察到主动脉瓣结构出现时,更换防脱玻片贴片,正式保留切片进行染色。冷冻切片干燥后入60%异丙醇浸洗10min ,油红O工作液染色10 min ,60%异丙醇分化3~10 s至间质清晰,水洗,Mayer苏木精复染1 min,自来水返蓝,用滤纸小心吸干水分,甘油明胶封片,Olympus倒置显微镜采集图像。
1.6 苏木精-伊红染色( hematoxylin-eosin,HE)取主动脉根部石蜡切片于60℃烤箱中30 min,迅速放入二甲苯中及100%、95%、80%、70%浓度梯度无水乙醇脱蜡处理后,迅速放入双蒸水中5 min,冲洗3次,苏木素染色15 min,冲洗后0.5%的盐酸乙醇3 s,冲洗后0.5%伊红染色2 min,依次70%、80%、95%、100%乙醇中脱水,二甲苯中固定后中性树胶封片。
1.7 免疫组化取主动脉根部石蜡切片于60℃烤箱中30 min,迅速放入二甲苯中及100%、95%、80%、70%浓度梯度无水乙醇脱蜡处理后,迅速放入双蒸水中5 min,冲洗3次,将玻片放入柠檬酸盐缓冲液中,微波炉中高火3 min,低火微沸10 min,进行抗原修复,自然冷却至室温,PBS冲洗。用3%的H2O2去离子水孵育10 min,以阻断内源性过氧化物酶,PBS冲洗,加入ST6Gal-Ⅰ一抗(1 ∶40稀释),4℃孵育过夜,PBS冲洗3次,滴加polymer helper室温孵育20 min,PBS冲洗,滴加poly-HRP anti-Rabbit IgG室温孵育20 min,PBS冲洗,DAB染色,苏木精复染,于70%、80%、95%、100%乙醇中脱水,二甲苯固定,中性树胶封片。
1.8 统计学分析 采用Graphpad Prism5.01软件进行数据分析,结果用 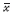 ± s表示。组间采用单因素方差分析及t检验。
± s表示。组间采用单因素方差分析及t检验。
在实验过程中小鼠均无异常死亡,基线模型组(baseline)、空白对照组(control)和瑞舒伐他汀组(rosuvastatin)造模前后的体重及血糖变化差异均无显著性,且3组小鼠的摄食量及重要脏器(心、肝、脾、肺、肾)的质量差异均无统计学意义。
2.1 瑞舒伐他汀对小鼠各种血脂水平的影响血脂检测结果显示,基线模型组小鼠血清中LDL-C、HDL-C、TC、TG均高于正常值,表明模型构建成功。23周高脂饲料饲养的对照组小鼠血清中LDL-C、TG和TC水平均高于基线模型组(Fig 1A~D)。但药物组小鼠血清中上述三指标均有所降低,表明瑞舒伐他汀的确具有降脂活性。

|
| Fig 1 Comparison of serum lipid levels in baseline group,control group and rosuvastatin group in ApoE-/- mice *P < 0.05 vs control |
经高脂喂养23周的对照组与16周的基线模型组比较,小鼠主动脉管弓的动脉斑块面积明显增加(P < 0.05)。而药物组与对照组比较,斑块面积明显减少(P < 0.05),见Fig 2。说明瑞舒伐他汀具有预防斑块形成的作用。

|
| Fig 2 Comparision of atherosclerosis plaquesin three groups in ApoE-/- mice *P < 0.05 vs control |
分析血管内膜的变化情况,结果如Fig 3所示,可见基线模型组小鼠血管主要病理改变为:内膜明显增厚,内皮排列紊乱且不连续,管壁厚度增加,平滑肌迁移内膜,中膜平滑肌明显萎缩变薄。表明模型构建成功。增加高脂喂养时间的对照组与基线模型组比较,病变呈明显的进行性加重趋势。动脉内膜连续性更弱,内膜厚度进一步增加,差异有显著性(P < 0.05)。瑞舒伐他汀药物组与对照组相比,内膜厚度明显降低(P < 0.05)。结果与上述油红O染色斑块面积变化趋势一致。

|
| Fig 3 Comparative analysis of atherosclerosisin aortic wall in ApoE-/- mice in three groups *P < 0.05 vs control |
为分析瑞舒伐他汀药理作用是否与唾液酸转移酶表达存在关联。分别取3组小鼠主动脉弓,制备石蜡切片,通过免疫组化分析唾液酸转移酶表达分布情况。结果显示,ST6Gal-Ⅰ在各组均有表达,3组血管内皮细胞层均可见棕色颗粒或棕色弧线。表明ST6Gal-Ⅰ主要分布于内膜下内皮细胞层。表明ST6Gal-Ⅰ主要分布于内膜、中膜平滑肌层。如Fig 4所示,对照组与基线模型组比较,血管内皮层ST6Gal-Ⅰ的表达量随着高脂饲料的长期喂养而明显降低。而瑞舒伐他汀药物组与对照组相比,随着AS的病理性变化减弱,血管内皮中ST6Gal-Ⅰ蛋白表达量有所增加。提示在AS发生、发展的进程中,α2,6-唾液酸转移酶的表达与之密切相关,且瑞舒伐他汀可以有效回复唾液酸转移酶的表达。瑞舒伐他汀可能通过影响糖基转移酶的表达来发挥治疗AS的作用。α2,6-唾液酸转移酶可能是个全新的药物作用靶点。

|
| Fig 4 Expression of ST6Gal-Ⅰ in aortic rootin ApoE-/- mice in three groups by immunohistochemistry |
动脉粥样硬化是心血管疾病中最常见的疾病,也是危害人类健康的常见病,已成为全球关注的热点。其形成基础是脂质代谢异常导致血管内皮细胞的损伤和功能紊乱,血管中膜平滑肌细胞表型发生转换、由内膜向中膜迁移、增殖并形成纤维帽以包裹泡沫细胞等粥样物质[15],诱发慢性炎症反应[3],从而形成动脉粥样斑块。瑞舒伐他汀因有较好的降脂及抗炎作用,可逆转粥样斑块形成,因此常用于抗动脉粥样硬化研究。近年来大量研究证实,瑞舒伐他汀除了调血脂作用外,还可通过调节内皮功能、抗血栓形成、稳定斑块、抑制血管炎症等多种途径治疗和预防AS 的发生发展。
唾液酸是9碳单糖,通常以短链残基的形式链接在糖蛋白、糖脂等糖缀合物的末端,如以α-2,3糖苷键连接到半乳糖(Gal)上,α-2,6糖苷键连接到半乳糖或N-乙酰半乳糖胺(GalNAc)上,α-2,8糖苷键连接到其他唾液酸上。唾液酸是重要的生物信息传递分子,细胞表面糖蛋白和糖脂的唾液酸化修饰在许多生物过程中发挥着至关重要的作用,包括细胞黏附、抗原识别和信号传导等。唾液酸糖基转移酶可以把胞苷一磷酸-β-N-乙酰神经氨酸(CMP-β-N-acetylneuramic acid)中的唾液酸基转移到糖蛋白和糖脂的链端,根据转移唾液酸基后新形成的糖苷键的类型可以把唾液酸糖基转移酶分为四类,其中ST6Gal-Ⅰ催化α-2,6糖苷键连接类型的糖蛋白形成,具有调节细胞之间的相互作用的潜力。如在结肠直肠癌转移灶中ST6Gal-Ⅰ的活性升高明显,而在正常结肠黏膜细胞中ST6Gal-Ⅰ的活性很低或不存在[16, 17, 18]。表明与肿瘤细胞转移关系密切。
本研究采用高脂饲养ApoE-/-小鼠构建AS模型,通过血液指标和组织形态分析,显示瑞舒伐他汀干预7周后,小鼠血脂水平与对照组比较,LDL-C、TG均明显下降(P < 0.05);油红O染色结果显示,瑞舒伐他汀干预组主动脉弓部斑块面积较对照组明显减小;HE染色亦显示瑞舒伐他汀组内膜厚度较对照组明显变薄,3组结果均说明瑞舒伐他汀具有降脂及抑制斑块形成的作用。免疫组化技术分析表明,对照组主动脉弓部管腔内皮层中ST6Gal-Ⅰ的表达明显降低。给予瑞舒伐他汀处理后,ST6Gal-Ⅰ的表达上调,说明动脉粥样硬化斑块的发展与唾液酸转移酶有密切关联。以往研究显示,AS是在各种炎症作用下发生的一系列病理改变,而炎症与糖基转移酶的变化密切相关。Doring等[12]发现在敲除ST3Gal-Ⅳ的小鼠中动脉粥样硬化的发生率明显较低。Gracheva等[19, 20]研究显示动脉粥样硬化病人动脉内膜下和血清中唾液酸糖基转移酶活性均明显增加。基于上述研究,我们认为糖基转移酶与动脉粥样硬化病理过程存在的密切联系是值得关注的,ST6Gal-Ⅰ表达变化可能是瑞舒伐他汀抑制动脉粥样硬化斑块形成的机制之一。因此,进一步揭示唾液酸转移酶与治疗动脉粥样硬化的分子机制,可以为临床治疗动脉粥样硬化提供新的靶点,但是更深层的作用机制还需进一步的研究。
(致谢:本实验在重庆医科大学生命科学研究院转化医学实验室完成,感谢重庆医科大学提供的科研平台,感谢实验主要参与人员张军老师、蒲强红师兄、邓潇师兄的帮助,感谢导师于超教授对该实验及论文撰写的悉心指导。)
| [1] | van Gils J M, Ramkhelawon B, Fernandes L, et al.Endothelial expression of guidance cues in vessel wall homeostasis dysregulation under proatherosclerotic conditions[J].Arterioscler Thromb Vasc Biol, 2013, 33(5):911-19. |
| [2] | Kriszbacher I, Koppan M, Bodis J.Inflammation, atherosclerosis, and coronary artery disease[J].N Engl J Med, 2005, 353(4):429-30. |
| [3] | Ross R.Atherosclerosis-an inflammatory disease[J].N Engl J Med, 1999, 340(2):115-26. |
| [4] | Gregersen I, Holm S, Dahl T B, et al.A focus on inflammation as a major risk factor for atherosclerotic cardiovascular diseases[J].Expert Rev Cardiovasc Ther, 2016, 14(3):391-403. |
| [5] | Ross R, Glomset J A.The pathogenesis of atherosclerosis(first of two parts)[J].N Engl J Med, 1976, 295(7):369-77. |
| [6] | Ross R, Glomset J A.The pathogenesis of atherosclerosis (second of two parts)[J].N Engl J Med, 1976, 295(8):420-25. |
| [7] | Lusis A J.Atherosclerosis[J].Nature, 2000, 407(6801):233-41. |
| [8] | 琚娜娜, 吴明军, 赵德璋, 等.白介素-1β通过上调血管内皮细胞中FucT-Ⅶ及蛋白糖基化调控细胞间黏附作用[J].中国药理学通报, 2013, 29(3):337-42. Ju N N, Wu M J, Zhao D Z, et al.Interleukin-1β regulates cell adhesion via up-reulating FucT-Ⅶ and proteins glycolation in vascular endothelial cells[J].Chin Pharmacol Bull, 2013, 29(3):337-42. |
| [9] | Zhuo Y, Bellis S L.Emerging role of alpha 2, 6-sialic acid as a negative regulator of galectin binding and function[J].J Biol Chem, 2011, 286(8):5935-41. |
| [10] | Woodard-Grice A V, McBrayer A C, Wakefield J K, et al.Proteolytic shedding of ST6Gal-Ⅰ by BACE1 regulates the glycosylation and function of alpha4beta1 integrins[J].J Biol Chem, 2008, 283(39):26364-73. |
| [11] | Sacco R L, Liao J K.Drug Insight:statins and stroke[J].Nat Clin Pract Cardiovasc Med, 2005, 2(11):576-84. |
| [12] | Doring Y, Noels H, Mandl M, et al.Deficiency of the sialyltransferase St3Gal4 reduces Ccl5-mediated myeloid cell recruitment and arrest:short communication[J].Circ Res, 2014, 114(6):976-81. |
| [13] | Yasukawa Z, Sato C, Kitajima K.Inflammation-dependent changes in alpha 2, 3-, alpha 2, 6-, and alpha 2, 8-sialic acid glycotopes on serum glycoproteins in mice[J].Glycobiology, 2005, 15(9):827-37. |
| [14] | Feig J E, Shang Y, Rotllan N, et al.Statins promote the regression of atherosclerosis via activation of the CCR7-dependent emigration pathway in macrophages[J].Plos One, 2011, 6(12):e28534. |
| [15] | Tedgui A, Mallat Z.Apoptosis as a determinant of atherothrombosis[J].Thromb Haemost, 2001, 86(1):420-26. |
| [16] | Le Marer N, Stehelin D.High alpha-2, 6-sialylation of N-acetyllactosamine sequences in ras-transformed rat fibroblasts correlates with high invasive potential[J].Glycobiology, 1995, 5(2):219-26. |
| [17] | Ito H, Hiraiwa N, Sawada-Kasugai M, et al.Altered mRNA expression of specific molecular species of fucosyl-and sialyl-transferases in human colorectal cancer tissues[J].Int J Cancer, 1997, 71(4):556-64. |
| [18] | Petretti T, Kemmner W, Schulze B, et al.Altered mRNA expression of glycosyltransferases in human colorectal carcinomas and liver metastases[J].GUT, 2000, 46(3):359-66. |
| [19] | Gracheva E V, Samovilova N N, Golovanova N K, et al.Sialyltransferase activity of human plasma and aortic intima is enhanced in atherosclerosis[J].Biochim Biophys Acta, 2002, 1586(1):123-28. |
| [20] | Gracheva E V, Samovilova N N, Golovanova N K, et al.Sialyltransferase activity in normal and atherosclerotic human aorta intima[J].Biochemistry(Mosc), 2001, 66(4):397-401. |