



2. 安徽医科大学第一附属医院药剂科, 安徽 合肥 230032
2. Dept of Pharmacy, the First Affiliated Hospital of Anhui Medical University, Hefei 230032, China
全反式维甲酸可诱导急性早幼粒细胞白血病(acute promyelocytic leukemia,APL)细胞分化成熟,疗效明显,其基本特点在于不直接杀伤肿瘤细胞而是诱导其分化为正常或接近正常的细胞。但由于ATRA会出现维甲酸综合症[1]、耐药性[2]等问题使其在临床上的应用受到了限制。本课题组以ATRA为先导化合物,通过对其碳链末端极性基团进行结构修饰,合成并经过体外药效学筛选发现4-氨基-2-三氟甲基苯基维甲酸酯(ATPR)具有较强的抑制肿瘤细胞增殖并诱导其分化的活性[3, 4, 5, 6, 7]。目前,ATPR已经申请自主知识产权(专利号: CN101591280、CN102008443A),有望成为一种新型抗肿瘤药物,为肿瘤患者带来希望。
蛋白质组(proteome)的概念最先由Marc Wilkins在1994年提出的,是指由一个基因组,或一个细胞、组织表达的所有蛋白质。近年来蛋白质组学技术被广泛应用到生命科学领域,并已成为寻找疾病分子标志和药物靶点最有效的方法之一。本研究以人白血病K562细胞为研究对象,通过蛋白质组学的方法,研究ATPR与ATRA分别作用于K562细胞后蛋白质的表达变化情况,并通过生物信息学分析来解释ATPR对K562细胞作用的特异性可能机制及分子靶点。
1 材料与方法 1.1 材料4-氨基-2-三氟甲基苯基维甲酸酯(Fig 1)由安徽医科大学药学院合成,纯度为99.66%,用无水乙醇溶解成浓度为20 mmol·L-1的贮存液,-20℃避光保存。K562细胞株购自中科院上海细胞库。RPMI 1640为Hyclone公司产品,小牛血清为杭州四季青公司产品。ATRA、二硫苏糖醇、碘乙酰胺为美国Sigma公司产品;总蛋白提取试剂盒购自Vazyme公司;蛋白纯化试剂盒(2D Clean-Up Kit)、蛋白定量试剂盒(2D Quant Kit)购自GE healthcare;Trypsin Gold(货号V5280)购自美国Promega公司;丙酮、乙腈、醋酸、甲醇、甲酸、三氟乙酸均为色谱级。高速冷冻离心机购自Beckman公司;ACCELA 600 Pump液相色谱和傅立叶变换静电场轨道肼质谱(LTQ Orbitrap XL)购自美国Thermo fisher公司; Xcalibur 2.1软件和Proteome Discoverer软件购自美国Thermo公司。

|
| Fig 1 4-amino-2-trifluoromethyl-phenyl retinate structural formula |
K562细胞采用含10%胎牛血清的RPMI 1640并加入青霉素1×105 IU·L-1和链霉素100 mg·L-1的培养体系培养。培养环境为37 ℃、5%的CO2培养箱,每2天更换1次细胞培养液,取对数生长期的细胞进行实验。随机分为对照组、ATPR及ATRA组,ATPR和ATRA的终浓度均为1×10-6 mol·L-1,培养箱中孵育48 h后收集细胞。
1.3 制备蛋白质组学样品离心收集细胞,加入500 μL细胞裂解液(临用前加入2 μL蛋白酶抑制剂),摇床震荡15 min后,离心15 min,吸出上清液即为细胞总蛋白。蛋白定量后取含200 μg总蛋白的裂解液,加入4倍体积的丙酮过夜沉淀。离心收集沉淀后加入12.5 μL 复溶液(尿素 7 mol,硫脲 2 mol,3-[(3-胆固醇氨丙基)二甲基氨基]-1-丙磺酸 65 mmol),完全溶解后加入87.5 μL 50 mmol 碳酸氢铵混匀。稀释后的蛋白溶解液加入4 μL 100 mmol的二硫苏糖醇,金属浴50℃ 15 min;避光,加入4 μL 300 mmol的碘乙酰胺,放置15 min,加入4 μg 胰酶(酶 ∶蛋白=1 ∶50)混匀,于37℃恒温水浴,孵育过夜。1 500×g 离心2 min,收集液体。使用Strata-X柱树脂脱盐后收集洗脱液,于旋转蒸发仪中干燥样品。
1.4 高效液相色谱-串联质谱(ESI-LC-MS/MS)分析干燥后的样品用30 μL 0.1%甲酸溶解后进入ESI-LC-MS/MS检测,流动相A为0.1%甲酸水溶液,流动相B为100%乙腈溶液,180 min梯度洗脱样品,过程为0~10 min,B相为10%;10~110 min,B相从10%上升至40%;110~140 min,B相从40%上升至95%;140~165 min,B相维持95%;165~170 min,A相从5%上升至90%;170~180 min,A相维持90%。洗脱的肽段经纳升级电喷雾离子源接口喷出,进入LTQ Orbitrap XL高分辨率质谱仪后分析。质谱扫描条件如下:采用数据依赖模式,在正离子模式下进行扫描,电喷雾电压:3.8 kV;质量扫描范围:300~2 000;离子传输毛细管温度为200℃;一级质谱Orbitrap分辨率:60 000;母离子窗口:2 u;串级质谱碰撞归一化能量:35。
1.5 蛋白质鉴定和生物信息学分析对质谱输出的原始文件通过Proteome Discovery 1.2 软件进行Sequest搜索,数据库为人源的蛋白序列数据库,来源于ftp://ftp.uniprot.org/pub/databases/uniprot/current_release/knowledgebase/proteomes/。检索参数如下:M/Z为350~5 000 u,母离子容忍度为10 ppm,碎片离子容忍度为0.8 u,肽段和蛋白质鉴定FDR均小于1%。利用生物信息学DAVID、KEGG、STRING数据库,分析鉴定出的差异蛋白质所具有的分子功能、所参与的生物学过程等信息。
1.6 统计学分析实验结果用 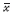 ± s表示,采用SPSS 16.0软件分析结果。组间数据单因素方差分析后采用LSD方法两两比较。
± s表示,采用SPSS 16.0软件分析结果。组间数据单因素方差分析后采用LSD方法两两比较。
毛细管色谱柱分离样品(Fig 2),质谱鉴定显示ATPR组共计542个蛋白质,ATRA组共计565个蛋白质。其中,ATPR组特异性的蛋白质有120个,ATRA组特异性的蛋白质有143个,ATPR和ATRA共有的蛋白质有422个(Fig 3)。

|
| Fig 2 The total ion current of groups A: ATPR group; B:ATRA group |

|
| Fig 3 Number of specific proteins in ATPR or ATRA groups and their common proteins |
使用DAVID数据库分析ATPR、ATRA各自的特异性蛋白质所参与的生物进程(Fig 4),ATPR特异性蛋白质参与39个生物学过程,主要有蛋白质和大分子的代谢、蛋白质转运和定位、大分子和蛋白复合物组装、翻译、RNA剪接、氮化合物生物合成、核苷酸生物合成等生物学过程,ATRA特异性蛋白质参与了22个生物学过程,涉及到翻译、激素代谢、9-顺式维甲酸代谢过程、蛋白质折叠、核糖体小亚基的生物合成、细胞蛋白质代谢过程的负调控、蛋白复合物组装、非编码RNA代谢等过程。

|
| Fig 4 The biological processes involvedin ATPR(A)or ATRA(B)specific proteins |
同时,使用KEGG pathway对特异性蛋白质可能参与的信号通路进行分析发现,ATPR组蛋白质主要参与Metabolic pathways、Ribosome、RNA transport、Spliceosome、Protein processing in endoplasmic reticulum、Purine metabolism、Parkinson’s disease、Proteoglycans in cancer、Pathways in cancer、PI3K-Akt signaling pathway、TGF-beta signaling pathway等信号通路,而ATRA组蛋白质则主要参与Ribosome、Metabolic pathways、RNA transport、Alzheimer’s disease、DNA replication、MAPK signaling pathway等信号通路。
2.3 ATPR组特异性蛋白质的相互作用网络分析通过STRING在线工具分析ATPR特异性蛋白质间的蛋白-蛋白相互作用网络(Fig 5),结果显示整个网络里,同其他≥6 个蛋白存在相互作用关系的至少有33个蛋白,其中以核糖体家族蛋白RPL3、RPL8、RPL13、RPL7A、RPL21、RPS3、RPS14、初期多肽相关复合体α亚基蛋白NACA、转录因子BTF3、真核翻译起始因子家族蛋白EIF3A、EIF6、非组蛋白染色体蛋白NHP2L1、蛋白质磷酸酶PPP2CA蛋白与其他≥10 个相关蛋白存在直接相互作用关系,为此蛋白相互作用网络的中心蛋白,删除这些中心蛋白后,互作网络结构就变得比较涣散。

|
| Fig 5 Protein-protein interaction map of ATPR group Coloured lines represent different evidences for each interaction: red line, gene fusion; green line, neighbourhood; blue line, co-occurrence; purple line, experimental evidences; yellow line, textmining; light blue line, databases; black line, co-expression. Bold numbers associated to proteins correspond to the different biological processes having statistical significance (P-value < 0.05) |
ATRA能够明显地抑制肿瘤细胞增殖并诱导其分化。但由于ATRA出现维甲酸综合症等问题,限制了其应用。课题组对ATRA进行结构修饰,合成了新型维甲酸衍生物ATPR。大量体内外研究表明[4, 5, 6, 7],新型维甲酸衍生物ATPR对多种肿瘤细胞具有一定的抑制增殖和诱导分化作用。前期研究[4]发现ATPR对K562细胞也具有抑制增殖和诱导分化作用。本研究通过对ATPR和ATRA分别作用于K562细胞后产生的蛋白质表达谱进行鉴定以及生物信息学分析,从整体上研究ATPR及ATRA对K562细胞作用下蛋白质组学变化中的差异,研究ATPR特异性作用于K562细胞的分子机制和靶点。
DAVID数据库分析结果说明,ATPR特异性蛋白质参与的生物学过程主要集中在蛋白质的组装、翻译、转运和代谢,核苷酸的生物合成等方面,与ATRA特异性蛋白质涉及的过程存在差异性。同时,KEGG结果进一步证实ATPR组特异性蛋白质主要参与蛋白质处理,癌症相关等通路,而ATRA组蛋白质则主要参与DNA复制和RNA转运等通路。通过STRING数据库分析ATPR特异性蛋白质的相互作用网络,发现以EIF3A、EIF6、RPL3、RPL8、RPL13、RPL7A、RPL21、RPS3、RPS14、NACA、BTF3等蛋白为中心构成相互作用的网络。
真核翻译起始因子(eukaryotic initiation factor ,EIF)是参与真核生物翻译过程的重要蛋白超家族。许多研究[8]表明,真核启动因子 EIF3A不仅在蛋白质翻译起始过程中起重要调控作用,还参与了细胞周期的调控,它与细胞的生长、分化和增殖存在着重要联系。EIF3A可以调控核糖核苷酸还原酶M2亚基(ribo nucleotide reductase M2 ,RRM2)和周期依赖性激酶抑制因子p27kip(cyclin dependent kinase inhibitor p27 ,p27kip)等因子。Liu 等[9]证实,EIF3A 对鼠结肠上皮细胞的分化发挥着重要的作用。这说明,EIF3A 在不成熟的细胞中有表达,并且其功能很可能是促进这些细胞向成熟阶段分化。另外,EIF3A与肿瘤的发生、发展及预后紧密相关。何晖等[10]认为EIF3A 的表达可能与肿瘤细胞的进展过程负相关;对于化疗药物干预的肿瘤细胞,EIF3A的高表达使肿瘤细胞不易产生耐药,患者的预后更好。之后的研究中发现分化程度高的肿瘤中,EIF3A的表达高于低分化的肿瘤组织,此外,EIF3A 高表达的患者减少了肿瘤转移。根据本课题组前期研究[4],ATPR可使细胞周期蛋白cyclin D1、cyclin E表达量减少,周期蛋白依赖性激酶CDK2、CDK4、CDK6也有不同程度的减少,细胞周期依赖激酶抑制因子p27kip1表达变化不明显而抑制因子p57kip2的表达量则明显增加,提示ATPR很可能是通过EIF3A的表达调控细胞周期,并诱导细胞分化,EIF3A可能是ATPR抑制K562细胞增殖并诱导其分化的靶点之一。
真核启动因子6(EIF6)又叫ITGB4BP、p27BBP,作为一种核基质,可转移至细胞核中调控翻译起始。Wnt信号通路调控细胞增殖和迁移、分化、凋亡等过程,其中β-catenin是关键分子。Wnt蛋白与Wnt配体结合后,会促使胞质内游离出β-catenin进入核内,与T细胞因子/淋巴细胞增强因子(T cell factor/lymphoid enhancer factor,TCF/LEF)结合,进而启动下游靶基因或蛋白(如c-myc、cyclin D1)的表达。一旦Wnt信号通路激活,可促进细胞增殖而不进入分化。EIF6可以明显下调Wnt/β-catenin信号通路的活性[11]。Wang等[12]发现EIF6高表达于分化后的细胞,提示可能参与癌细胞分化。结合本课题组前期研究ATPR诱导NB4细胞分化过程中[13],c-myc蛋白表达与端粒酶逆转录酶hTERT的表达水平均呈下降趋势,所以预测ATPR可能是通过EIF6表达来抑制c-myc蛋白从而抑制hTERT及端粒酶活性,最终诱导NB4细胞分化,提示EIF6可能是ATPR作用于K562细胞的靶点。
已有研究表明[14],核糖体蛋白不仅参与蛋白质合成,而且还参与其他功能,如细胞生长、分化、凋亡等。核糖体蛋白( ribosomal protein)表达下调及功能不全可导致肿瘤的发生。作为核糖体大亚基蛋白,核糖体蛋白RPL3可通过p53来调控细胞周期和凋亡,RPL3可致细胞G1/S期阻滞或细胞凋亡[15]。核糖体蛋白RPL8已被发现在乳腺癌、黑色素瘤等肿瘤中表达[16]。核糖体蛋白RPL13参与调控抑癌基因等过程[17]。核糖体蛋白RPL7A则与细胞的生长、发育、分化有关,且其所在9号染色体往往在骨肉瘤中缺失[14],Burris等[18]报道,RPL7A还能特异性结合维甲酸受体(retinoic acid receptor,RAR)。而作为核糖体小亚基蛋白,在临床上,RPS3的异常表达与白血病、乳腺癌、肺癌等相关。郭恒西等[19]研究显示下调RPS3表达,会造成细胞G2/M期及S期阻滞等。另外,Ebert等[20]报道,减少造血干细胞内核糖体小亚基蛋白RPS14表达,可导致造血干细胞向红系分化受阻。本研究结果表明,ATPR作用后,可诱导K562细胞核糖体大小亚基蛋白RPL3、RPL8、RPL13、RPL7A、RPL21、RPS3、RPS14表达,而在ATRA组没有诱导这些蛋白表达,提示这些核糖体亚基蛋白也可能是ATPR特异性发挥作用的靶标。
初期多肽相关复合体(nascent polypeptide-associated complex,NAC)是一种多功能的蛋白,其α亚基即NACA,能调控Fas相关死亡域蛋白(Fas associated with death domain protein ,FADD)等一系列相关的功能。Lopez等[21]研究了NACA在人体造血中的作用,发现NACA不仅能加快红细胞的分化速度,还可以正向调控人红细胞的分化。与NACA蛋白二聚化的对象为转录因子BTF3。Liu等[22]在胃癌SGC7901细胞中发现BTF3可使细胞发生G1期阻滞,下调BTF3导致细胞凋亡。还有研究[23]证实了蛋白质磷酸酶PPP2CA参与皮肤组织分化信号通路。
综上所述,本研究结果表明,ATPR诱导的特异性中心蛋白质的改变均参与对细胞生长增殖、诱导细胞分化和凋亡等过程的调控,而ATRA作用下不具有这些蛋白质,这是两者的差异性。另外,我们推断,这些特异性蛋白质及其组建的相互作用网络,是ATPR抑制K562细胞增殖并诱导其分化等作用的可能机制,而本研究也从整体上为ATPR诱导K562细胞分化作用的分子靶点提供理论基础。
(致谢:本实验在安徽医科大学实验室完成,衷心感谢导师陈飞虎教授对我的课题的悉心指导,感谢夏泉老师对我的课题及实验的耐心传授,感谢葛金芳老师课题的细心解答,感谢实验室的同学们对我的实验提供的帮助。)
| [1] | Dhar A K, Barman P K. Retinoic acid proteomics syndrome-cardiac complication[J].J Assoc Physicians India, 2012, 60:63-5. |
| [2] | Tomita A, Kiyoi H, Naoe T. Mechanisms of action and resistance to all-trans retinoic acid (ATRA) and arsenic trioxide(As2O3) in acute promyelocytic leukemia[J].Int J Hematol, 2013, 97(6):717-25. |
| [3] | 阮晶晶, 陈飞虎, 徐佼, 等. 新型维甲酸衍生物诱导NB4细胞分化的初步研究[J].中国临床药理学与治疗学, 2008, 13(11):1237-42. Ruan J J, Chen F H, Xu J, et al. Preliminary study of NB4 cell differentiation induced by retinoic acid derivatives[J].Chin J Clin Pharmacol Ther, 2008, 13(11):1237-42. |
| [4] | 阮晶晶, 陈飞虎, 徐佼, 等. 4-氨基-2-三氟甲基苯基维甲酸酯对K562细胞分化和细胞周期的影响[J]. 中国药理学通报, 2009, 25(9):1238-43. Ruan J J, Chen F H, Xu J, et al. 4-amino-2-trifluoromethyl-phenyl retinate induced K562 cell differentiation and cell cycle influence[J]. Chin Pharmacol Bull, 2009, 25(9):1238-43. |
| [5] | 汪楠, 陈飞虎, 葛金芳, 等. 4-氨基-2-三氟甲基苯基维甲酸酯对MCF-7细胞增殖和分化的影响及其机制研究[J]. 中国药理学通报, 2013, 29(6):767-72. Wang N, Chen F H, Ge J F, et al. Effects of 4-amino-2-trifluoromethyl-phenyl retinate on proliferation and differentiation of MCF-7 cell and its possible mechanisms[J]. Chin Pharmacol Bull, 2013, 29(6):767-72. |
| [6] | 彭晓清, 陈飞虎, 葛金芳, 等. 4-氨基-2-三氟甲基苯基维甲酸酯通过PTEN/PI3K/Akt抑制YAC-1细胞增殖和诱导其分化[J]. 中国药理学通报, 2014, 30(1):60-5. Peng X Q, Chen F H, Ge J F, et al. Effects of 4-amino-2-trifluoromethyl-phenyl retinate on proliferation and differentiation of YAC-1 cells via PTEN/PI3K/Akt[J]. Chin Pharmacol Bull, 2014, 30(1):60-5. |
| [7] | 雷静, 陈飞虎, 葛金芳, 等. 4-氨基-2-三氟甲基苯基维甲酸酯对人乳腺癌细胞株MDA-MB-231诱导分化作用及可能的机制研究[J].中国药理学通报, 2015, 31(7):973-9. Lei J, Chen F H, Ge J F, et al. Inducing effect of 4-amino-2-trifluoromethyl-phenyl retinate on differentiation of human breast cancer MDA-MB-231 cell and its possible mechanisms[J].Chin Pharmacol Bull, 2015, 31(7):973-9. |
| [8] | 徐潇静, 周宏灏, 刘昭前. eIF3a的生物学功能研究进展[J]. 中国临床药理学与治疗学, 2010, 15(2):219-24. Xu X J, Zhou H H, Liu Z Q. Recent research progress of eIF3 in biological function[J].Chin J Clin Pharmacol Ther, 2010, 15(2):219-24. |
| [9] | Liu Z, Dong Z, Yang Z, et al. Role of eIF3a (eIF3 p170) in intestinal cell differentiation and its association with early development[J]. Differentiation, 2007, 75(7):652-61. |
| [10] | 何晖, 尹继业, 周宏灏, 等. eIF3a与肿瘤的研究进展[J]. 中国药理学通报, 2014,30(4):445-8. He H, Yin J Y, Zhou H H, et al. Research progress on eIF3a and cancer[J].Chin Pharmacol Bull, 2014, 30(4):445-8. |
| [11] | 赖庆梧, 彭旭, 谭江琳, 等. ITGB4BP对Wnt/β-catenin信号通路活性的下调作用[J]. 第三军医大学学报, 2010,32(18):1945-8. Lai Q W, Peng X, Tan J L, et al. ITGB4BP downregulates activity of Wnt/β-catenin signaling pathway in HEK293 cells[J]. J Third Mil Med Univ, 2010, 32(18):1945-8. |
| [12] | Wang J, Jiao H, Stewart T L, et al. Improvement in postburn hypertrophic scar after treatment with IFN-alpha2b is associated with decreased fibrocytes[J]. J Interferon Cytokine Res, 2007, 27(11):921-30. |
| [13] | 陈慧慧, 陈飞虎, 徐佼, 等. c-myc、hTERT在4-氨基-2-三氟甲基苯基维甲酸酯诱导NB4细胞分化中的作用[J]. 安徽医科大学学报, 2011, 46(3):235-9. Chen H H, Chen F H, Xu J, et al. Effects of c-myc, hTERT on differentiation of NB4 cells induced by 4-amino-2-trifluoromethy-l phenyl retinate[J].Acta Univ Med Anhui, 2011, 46(3):235-9. |
| [14] | 郑水儿, 林峰, 沈赞, 等. 核糖体蛋白L7A在骨肉瘤组织中的表达及临床意义[J].中国癌症杂志,2009, 19(2):101-5. Zheng S E, Lin F, Shen Z, et al. Expression of ribosomal protein L7a (RPL7A) in human osteosarcoma and its clinical significance[J]. China Oncol, 2009, 19(2):101-5. |
| [15] | Russo A, Esposito D, Catillo M, et al. Human rpL3 induces G1/S arrest or apoptosis by modulating p21waf1/cip1 levels in a p53-independent manner[J]. Cell Cycle, 2013, 12(1):76-87. |
| [16] | Swoboda R K, Somasundaram R, Caputo L, et al. Shared MHC class II-dependent melanoma ribosomal protein L8 identified by phage display[J].Cancer Res, 2007, 67(8):3555-9. |
| [17] | Takemasa I, Kittaka N, Hitora T, et al. Potential biological insights revealed by an integrated assessment of proteomic and transcriptomic data in human colorectal cancer[J].Int J Oncol, 2012, 40(2):551-9. |
| [18] | Burris T P, Nawaz Z, Tsai M J, et al. A nuclear hormone receptor-associated protein that inhibits transactivation by the thyroid hormone and retinoic acid receptors[J].Proc Natl Acad Sci USA, 1995, 92(21):9525-9. |
| [19] | 郭恒西, 陈慧, 詹轶群, 等. 核糖体蛋白RPS3 Ser209影响NF-κB转录活性及其与DNA结合能力[J]. 军事医学, 2015, 39(2):117-22. Guo H X, Chen H, Zhan Y Q, et al. Effect of ribosomal protein S3 Ser209 on NF-κB transcriptional activity and its DNA binding capacity[J].Mili Med Sci, 2015, 39(2):117-22. |
| [20] | Ebert B L, Pretz J, Bosco J, et al. Identification of RPS14 as a 5q-syndrome gene by RNA interference screen[J]. Nature, 2008, 451(7176):335-9. |
| [21] | Lopez S, Stuhl L, Fichelson S, et al. NACA is a positive regulator of human erythroid-cell differentiation[J]. J Cell Sci, 2005, 118(8):1595-605. |
| [22] | Liu Q, Zhou J P, Li B, et al. Basic transcription factor 3 is involved in gastric cancer development and progression[J]. World J Gastroenterol, 2013, 19(28):4495-503. |
| [23] | 陈霞. 小鼠皮肤特异性敲除Ppp2ca基因对毛发生长的影响[D]. 南京:南京医科大学, 2014. Chen X. Effect of mouse skin specific knockout Ppp2ca gene on hair growth[D]. Nanjing:Nanjing Medical University, 2014. |