


随着生活节奏的加快,社会压力的增加,抑郁症的发病率逐年升高,严重危害着人类的身心健康。抗抑郁药物的研发及筛选对于临床抑郁症的治疗有及其重要的意义,而稳定可靠的抑郁动物模型对抗抑郁药物的研发及筛选有非常重要的价值。嗅球切除是一种常用的抑郁研究模型,通过手术摘除或者破坏嗅球,直接导致动物嗅觉丧失和多个脑区神经元的退化。目前对于嗅球切除抑郁动物模型已有一定的研究,包括嗅球切除后动物的多种行为,如活动增强,被动逃避能力下降,学习记忆能力衰减,应激反应增强,同时进食和性行为发生改变等,这些症状与激越性抑郁症相类似[1]。从这些行为改变中恢复正常需要长期服用抗抑郁类药物,这与临床抗抑郁治疗的时程相似。尽管嗅球切除模型已经广为应用,并且对术后动物海马、前额叶、中脑、脑干等多个脑区神经递质及相关蛋白表达影响也有研究报道[2, 3, 4],但研究并不全面和系统,且很多报道结果不一,甚至存在相反的结论。本实验系统研究了嗅球切除对动物海马多种神经递质,包括5-羟色胺(5-hydroxytryptamine,5-HT)、5-羟吲哚乙酸(5-hydroxyindoleacetic acid,5-HIAA)、去甲肾上腺素(noradrenaline,NE)、多巴胺(dopamine,DA)、肾上腺素(epinephrine,Epi)、谷氨酸(glutamate,Glu)和γ氨基丁酸(γ-aminobutyric acid,GABA)等,以及相关受体、转运蛋白、神经再生和炎症相关蛋白基因表达的影响,同时研究了嗅球切除对C57BL/6小鼠抑郁及焦虑行为的作用,为该模型更好地应用于科研和抗抑郁药物的研发提供了参考。
1 材料 1.1 药品与试剂异氟烷购自河北九派制药股份有限公司(批号:130601),注射用青霉素钠购自华北制药股份有限公司(批号:H13020657),戊巴比妥钠(批号:WS20120912),色谱甲酸、色谱甲醇、乙腈购自赛默飞世尔科技(中国)有限公司。
1.2 实验动物♂5周龄C57BL/6小鼠购自上海斯莱克实验动物有限责任公司,并在上海中医医药大学动物实验中心(25±1)℃,40%~60%相对湿度,昼夜各12 h SPF动物房自由进食饲养,实验动物使用许可证号:SCXK(沪)2012-002
1.3 仪器液相质谱联用仪购自安捷伦(型号DEBAF01206),高架迷宫、旷场、悬尾及强迫游泳测试仪购自上海玉研,RMA-13-SSV小鼠麻醉机购自Medical Supplies & Services Int Ltd,高通量研磨仪由上海净信科技提供(批号TL48E),C18色谱柱(2.1×100 mm,1.7 μm)购自Waters。
2 方法 2.1 动物分组及手术建模C57BL/6小鼠自购买后,适应性饲养1周,随机分为假手术组(sham)和嗅球切除模型组(OB),每组12只动物。异氟烷麻醉小鼠后,在两耳连线中点处切开皮肤,暴露颅骨,电动磨钻于嗅球处中线两侧将颅骨各钻1个小孔,探针搅动破坏嗅球后,真空泵将破坏的嗅球组织吸出,吸收性明胶海绵填孔止血。假手术小鼠采用相同的手术方法,但在手术定位点上钻孔后不损伤嗅球。青霉素溶液(20万U·mL-1)冲洗手术切口,缝合皮肤,并肌注青霉素钠每只4万U,连续给药3 d,以防术后感染。
2.2 行为学检测小鼠手术后恢复18 d,分别进行如下行为学测定: ① 强迫游泳测试(FST),将小鼠置于预先注有20 cm深清水,温度(25±1) ℃的圆柱型透明器皿内(高30 cm,直径20 cm),分别记录每只小鼠在6 min内的累计不动时间,比较后4 min内的不动时间; ② 悬尾测试(TST),用胶带将小鼠尾部(距尾端2 cm)固定于一根水平木棍上,使动物呈倒挂状态,其头部离水平面5~6 cm,分别记录每只动物在6 min内后4 min的累计不动时间;③ 旷场测试(OFT),将小鼠放入开场试验箱正中间,软件记录5 min内小鼠总活动路程;④ 高架迷宫测试(EPMT) 将高架迷宫放在光线均匀的实验区域内,分别将小鼠放入迷宫中央部位,实验室开始时使头部面向开臂,记录5 min内小鼠进入开臂的时间和次数。
2.3 标本采集完成所有的行为学测试后,1%戊巴比妥钠麻醉小鼠,于冰上分离小鼠脑组织,取出海马后置EP管中,迅速放入液氮冷冻,并随后保存在-80℃冰箱备用。
2.4 神经递质检测海马组织称重后,每份加入700 μL的甲酸-甲醇(1 ∶ 1 000) 和10 μL (10 μg·L-1,3,4-dihydroxybenzylamine)内标,匀浆,23 000 r·min-1离心15 min(4℃)。取上清,氮气吹干,每份再加100 μL流动相溶解,23 000 r·min-1离心15 min(4℃)后,取上清用于进一步分析。 LC-MS/MS色谱条件参考本实验室已发表文献 [5],液相条件如下:以甲酸-水(1 ∶ 1 000,A相),乙腈(B相)为流动相进行梯度洗脱,0~4 min,0.02 B;6 min,0.8 B;8~10 min,0.9 B;流速为0.1 mL·min-1。
2.5 实时定量PCR分析取海马组织,TRIzol法提取总RNA,经DNaseI处理去除DNA污染后,试剂盒(Revert Aid First Strand Synthesis kit,Fermentas)反转录为cDNA,按照Taqman SYBR kit (Life Technologies)试剂盒说明方法进行实时定量PCR。PCR使用引物序列参见Tab1,样品的基因表达均以各自的甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)作为内参。

如Fig1A所示,切除嗅球之后,和假手术小鼠对比,模型组小鼠活动量明显增加,在旷场中的水平运动距离明显延长,差异有显著性(P<0.05)。与此相对应,在高架迷宫中(Fig1B),模型组小鼠进入开臂时间长于空白组,差异有显著性(P<0.01)。但在强迫游泳和悬尾测试中(Fig1C、1D),模型组小鼠的游泳不动时间与对照组比较,有上升的趋势,但差异无统计学意义。

|
Fig 1
Effect of olfactory bulbectomy on mouse behaviors(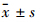 ,n=12)
A: Total distance in open-field test (OFT); B: Time spent in open arm of elevated plus maze test (EPMT); C: Immobility time in forced swimming test (FST); D: Immobility time in tail-suspension test (TST).*P<0.05, **P<0.01 vs Sham ,n=12)
A: Total distance in open-field test (OFT); B: Time spent in open arm of elevated plus maze test (EPMT); C: Immobility time in forced swimming test (FST); D: Immobility time in tail-suspension test (TST).*P<0.05, **P<0.01 vs Sham
|
如Tab2所示,嗅球切除改变了小鼠海马神经递质表达。结果显示,模型组小鼠海马5-HT含量明显上升,和假手术组比较差异具有显著性(P<0.01)。而5-HIAA含量在模型组中则明显下降(P<0.05)。与此相对应,5-HIAA与5-HT含量的比值也明显降低,差异有显著性(P<0.01)。尽管DA、Epi、NE及 GABA的含量在嗅球切除组小鼠的海马中没有变化,但Glu含量明显升高(P<0.05),而Glu与GABA的比值也有明显上升(P<0.01)。
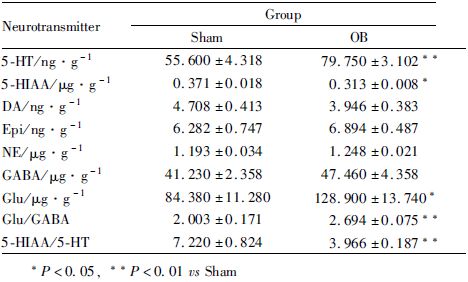
基于海马神经递质LC-MS/MS检测结果,我们进一步对5-HT、GABA以及Glu代谢、受体相关基因表达进行分析,同时对神经营养因子和炎症相关基因也做了检测。结果如Tab3所示,嗅球切除影响了海马中多 种基因的表达,TPH2 mRNA表达下调(P<0.05),BDNF/Trkb mRNA表达上调(P<0.05),TNF-α/CD11b mRNA表达上调(P<0.05),而其它基因则无显著变化。
,n=5)
 |
可靠稳定的抑郁动物模型对于抑郁症发病机制、抗抑郁药物的筛选及其抗抑郁作用机制研究有重要的意义。目前常用的抑郁动物模型有慢性轻度不可预见性应激模型、嗅球切除模型、行为绝望模型、习得性无助模型和一些药物诱导模型如:糖皮质激素诱导、LPS诱导、利血平逆转、5-羟色胺酸诱导大小鼠甩头模型等[6, 7, 8, 9, 10]。另外,转基因抑郁动物模型作为新型抑郁模型也得到了广泛的应用[11, 12]。然而,诸多模型都不能完全模拟临床抑郁病人的症状,只能针对部分病症研究,所以在现有研究基础上应考虑运用多种模型结合来扬长避短。
嗅球切除抑郁模型是常用模型之一,有研究报道,嗅球切除后,小鼠表现为旷场试验活动量增加,高架迷宫测试进入开臂时间增加[1],本实验研究结果与文献报道一致。然而,在强迫游泳和悬尾测试实验中,有研究结果表明嗅球切除后小鼠不动时间增加[13],与典型的抑郁模型结果一致,而有些研究结果却得出相反的结果,显示小鼠嗅球切除后游泳不动时间反而减少[2],本实验采用6周龄成年C57BL/6♂小鼠,手术18 d后行为学测试,尽量减少手术创伤对小鼠行为学的影响。结果显示,模型组较假手术组强迫游泳和悬尾不动时间均有一定程度的增加,但并没有显著性差异。分析其结果不一致的原因,可能与手术时小鼠是否成年、手术恢复后至行为学测试的时间长短、小鼠样本数以及各实验室具体的饲养和测试条件有关。
抑郁症与脑内神经递质5-HT及其代谢产物5-HIAA的含量密切相关。研究表明小鼠嗅球切除后5-HIAA/5-HT下降[14],表明5-HT的代谢功能下降。本实验对海马神经递质检测,结果为嗅球切除组小鼠海马内5-HT含量升高,5-HIAA/5-HT明显降低,海马内5-HT合成限速酶TPH2基因表达降低,则表明在5-HT合成降低的基础上5-HIAA与5-HT的比值也降低,可能是由于5-HT的代谢和转运功能降低,进而造成反馈抑制,影响了TPH2的基因表达。正常生理状态下,神经细胞间隙存在的兴奋性氨基酸与抑制性氨基酸维持平衡,当两者之间平衡失调,则会产生情绪紊乱如抑郁、焦虑[15]。本实验嗅球切除模型组小鼠海马内兴奋性氨基酸Glu与抑制性氨基酸GABA比值增高,表明细胞膜外堆积大量的兴奋性氨基酸,作用于膜受体后导致一系列变化,最终引起神经元的损伤,从而导致小鼠抑郁焦虑样表现。
TrkB为脑源性神经营养因子BDNF较高亲和力的受体,BDNF通过激活受体TrkB,对神经元的生存及活动起重要调节作用[16]。文献对嗅球切除后动物脑内BDNF变化的报道存在不一致,如Hellweg等[14]对嗅球切除16 d后小鼠海马及额前叶内BDNF检测结果显示其表达增高等,而Hendriksen 等[17]研究在SD大鼠嗅球切除10周后检测到海马内BDNF mRNA水平明显降低。也有实验对嗅球切除15 d后和20 d大鼠海马BDNF检测,发现其没有改变[18, 19]。本文对海马内BDNF mRNA及其受体TrkB mRNA的表达进行检测,发现二者表达同样升高,并且伴随着胶质细胞源性的神经营养因子GDNF的增高,但并没有伴随脑源性神经生长因子NGF的表达上调。对于嗅球切除后海马内BDNF结果报道不一致及本实验结果分析,其原因可能为种属的不同,在小鼠中,嗅球切除后会有一段适应性反应,在此时间段内,小鼠海马神经元反馈性调节脑源性神经营养因子,使其表达上调,有助于神经损伤的修复。
大量研究表明,抑郁症与炎症密切相关,抑郁症患者存在免疫功能激活[20]。TNF-α属于前炎症细胞因子,是一系列炎症反应中最早产生的细胞因子,临床研究表明抑郁患者脑内TNF-α处于激活状态[21]。CD11b是白细胞黏附分子β2整合素的a亚单位,与白细胞和内皮细胞的黏附有关,正常情况下,CD11b在中性粒细胞表面低水平表达,贮存在细胞的胞质颗粒中,在一系列前炎症调节因子,如LPS、TNF-α、IL-1β等刺激下,迅速易位至细胞膜,在细胞表面大量表达[22]。本实验发现小鼠海马内TNF-α和CD11b mRNA表达上调,则表明嗅球切除后小鼠免疫力下降导致炎症相关因子表达上调。
综上所述,本实验发现嗅球切除不仅影响了动物的行为,还影响了海马内多种神经递质及其相关代谢产物、神经营养因子、炎症相关基因的表达,提示抗抑郁药物的筛选及机制研究需综合应用多种动物模型并考虑多种因素影响,从而得出科学合理的结论。
| [1] | Song C, Leonard B E. The olfactory bulbectomised rat as a model of depression[J]. Neurosci Biobehav Rev, 2005, 29(4-5):627-47. |
| [2] | Mucignat-Caretta C, Bondí M, Caretta A. Time course of alterations after olfactory bulbectomy in mice[J]. Physiol Behav, 2006, 89(5):637-43. |
| [3] | Yuan T F, Slotnick B M. Roles of olfactory system dysfunction in depression[J]. Prog Neuropsychopharmacol Biol Psychiatry, 2014, 54:26-30. |
| [4] | Morales-Medina J C, Juarez I, Iannitti T, Flores G. Olfactory bulbectomy induces neuronal rearrangement in the entorhinal cortex in the rat[J]. J Chem Neuroanat, 2013, 52:80-6. |
| [5] | Huang F, Li J, Shi H L, et al. Simultaneous quantification of seven hippocampal neurotransmitters in depression mice by LC-MS/MS[J]. J Neurosci Meth, 2014, 229:8-14. |
| [6] | Ping G, Qian W, Song G, Zhaochun S. Valsartan reverses depressive/anxiety-like behavior and induces hippocampal neurogenesis and expression of BDNF protein in unpredictable chronic mild stress mice[J]. Pharmacol Biochem Behav, 2014, 124:5-12. |
| [7] | Pignatelli M, Vollmayr B, Richter S H, et al. Enhanced mGlu5-receptor dependent long-term depression at the Schaffer collateral-CA1 synapse of congenitally learned helpless rats[J]. Neuropharmacology, 2013, 66:339-47. |
| [8] | Zhao Y, Ma R, Shen J, et al. A mouse model of depression induced by repeated corticosterone injections[J]. Eur J Pharmacol, 2008, 581(1-2):113-20. |
| [9] | Kubera M, Curzytek K, Duda W, et al. A new animal model of (chronic) depression induced by repeated and intermittent lipopolysaccharide administration for 4 months[J]. Brain Behav Immun, 2013, 31:96-104. |
| [10] | Tian J, Zhang F, Cheng J, et al. Antidepressant-like activity of adhyperforin, a novel constituent of hypericum perforatum L[J]. Sci Rep, 2014, 4:5632. |
| [11] | Chen Y, Liu X, Jia X, et al. Anxiety-and depressive-like behaviors in olfactory deficient Cnga2 knockout mice[J]. Behav Brain Res, 2014, 275:219-24. |
| [12] | Xing B, Liu P, Jiang W H, et al. Effects of immobilization stress on emotional behaviors in dopamine D3 receptor knockout mice[J]. Behav Brain Res, 2013, 243:261-6. |
| [13] | Islam MR, Moriguchi S, Tagashira H, Fukunaga K. Rivastigmine improves hippocampal neurogenesis and depression-like behaviors via 5-HT1A receptor stimulation in olfactory bulbectomized mice[J]. Neuroscience, 2014, 272:116-30. |
| [14] | Hellweg R, Zueger M, Fink K, et al. Olfactory bulbectomy in mice leads to increased BDNF levels and decreased serotonin turnover in depression-related brain areas[J]. Neurobiol Dis, 2007, 25(1):1-7. |
| [15] | Sanacora G, Gueorguieva R, Epperson C N, et al. Subtype-specific alterations of gamma-aminobutyric acid and glutamate in patients with major depression[J]. Arch Gen Psychiatry, 2004, 61(7):705-13. |
| [16] | Numakawa T, Richards M, Nakajima S, et al. The role of brain-derived neurotrophic factor in comorbid depression:possible linkage with steroid hormones, cytokines, and nutrition[J]. Front Psychiatry, 2014, 5:136. |
| [17] | Hendriksen H, Meulendijks D, Douma T N, et al. Environmental enrichment has antidepressant-like action without improving learning and memory deficits in olfactory bulbectomized rats[J]. Neuropharmacology, 2012, 62(1):270-7. |
| [18] | Luo K R, Hong C J, Liou Y J, et al. Differential regulation of neurotrophin S100B and BDNF in two rat models of depression[J]. Prog Neuropsychopharmacol Biol Psychiatry, 2010, 34(8):1433-9. |
| [19] | Van Hoomissen J D, Chambliss H O, Holmes P V, et al. Effects of chronic exercise and imipramine on mRNA for BDNF after olfactory bulbectomy in rat[J]. Brain Res, 2003, 974(1-2):228-35. |
| [20] | Grudet C, Malm J, Westrin A, Brundin L. Suicidal patients are deficient in vitamin D, associated with a pro-inflammatory status in the blood[J]. Psychoneuroendocrinology, 2014, 50:210-9. |
| [21] | Shelton R C, Claibome J, Sidoryk-Wegrzynowicz M, et al. Altered expression of genesinvolved in inflammation and apoptosis in frontal cortex in major depression[J]. Mol Psychiatry, 2011, 16(7):751-62. |
| [22] | Borjesson D L, Simon S I, Hodzic E, et al. Kinetics of CD11b/CD18 up-regulation during infection with the agent of human granulocytic ehrlichiosis in mice[J]. Lab Invest, 2002, 82(3):303-11. |