



2. 南华大学肿瘤研究所, 湖南省高校肿瘤细胞与分子病理学重点实验室, 湖南 衡阳 421002;
3. 南华大学附属第一医院肿瘤科, 湖南 衡阳 421002
2. Cancer Research Institute, Key Laboratory of Cancer Cellular and Molecular Pathology of Hunan Provincial University, Hengyang Hunan 421002, China;
3. Dept of Oncology, the First Affiliated Hospital, University of South China, Hengyang Hunan 421002, China
白血病是常见恶性肿瘤之一,全世界每年新发病例约30万,死亡率高达70%,严重威胁着人类健康[1]。研究表明,二烯丙基二硫(diallyl disulfide,DADS)可抑制白血病等多种肿瘤增殖,其机制与增加组蛋白乙酰化修饰,阻滞肿瘤细胞周期,促进肿瘤细胞凋亡和诱导分化等有关[2]。我们前期工作证实,DADS能明显抑制人白血病HL-60细胞增殖和诱导凋亡,其机制与G2/M期阻滞,抑制ERK和激活p38、JNK信号通路,活化caspase,下调Bag-1、Bcl-2、PCNA、Mcl-1、CDK1,上调Fas-L、Bax,通过上调Rac2介导Rac2-NADPH氧化酶-ROS-JNK通路等有关[3, 4, 5, 6, 7, 8]。
肿瘤是多基因变化导致细胞周期紊乱的一类疾病,G2/M期检测点激酶Chk1和Chk2在细胞癌变过程中起着至关重要的作用,其介导Cdc25 磷酸酶与Cdks的活化和抑制是控制细胞G2/M期的关键[9]。本研究探讨DADS诱导HL-60细胞周期阻滞的分子机制,了解Chk与细胞周期之间的相互作用,从而为DADS防治HL-60细胞的信号干预治疗提供新思路。
1 材料与方法 1.1 细胞与试剂HL-60细胞为人急性髓细胞性白血病细胞株,由本实验室保存。DADS:Fluka公司产品,d420=1.0,Mr 146.28,纯度80%,含10%~20% DAS。DADS与Tween 80以1 ∶ 2的比例充分溶解后,加入体积分数为0.90的生理盐水稀释100倍,震荡混匀,作为母液保存于-20℃冰箱中,临用时将此母液稀释至所需浓度。Tween 80为华美公司产品。培养液:Gibco公司产品。RPMI 1640培养基10.4 g溶于1 000 mL蒸馏水中,加NaHCO3 2 g,用1 mol·L-1 HCl调pH值为7.0,抽滤除菌,4℃冰箱保存,临用时加10%小牛血清。小牛血清:杭州四季青生物工程公司产品,临用时经56℃水浴30 min灭活,-20℃保存。BCA蛋白定量检测试剂盒购自美国Pierce公司;显影定影试剂盒、丽春红染色液、预染蛋白Marker购于Fermentas公司。ECL发光检测试剂盒、Chk1(FL-467)、Chk2(H-300)、14-3-3 (aC-12)与CDK1多克隆抗体购于Santa Cruz公司;P-Chk1 (Ser345) 多克隆抗体购自Cell Signaling Technology公司;P-Chk2 (T387) 多克隆抗体购于美国Abzoom公司;羊抗兔二抗和羊抗鼠二抗购自KPL 公司。
1.2 细胞培养在37℃、饱和湿度、5%CO2孵箱中常规培养,培养液为含10%小牛血清的RPMI-1640,每2~3天换液传代,取对数生长期细胞用于实验。对照组不加DADS,溶媒对照组加等量的Tween 80,处理组加DADS。
1.3 细胞计数取对数生长期细胞,接种于24孔培养板,加处理因素,每24 h取4复孔细胞计数,每孔计数4次,求出3孔细胞的均值及标准差,连续计数4 d,绘制细胞生长曲线。肿瘤细胞群体倍增时间按公式计算:TD=t×lg2/lg(Nt/N0)(Nt为t时间的细胞数,N0为初种细胞数)。
1.4 软琼脂克隆形成实验取对数生长期细胞,离心,加培养基重悬使之分散成单个细胞,调整细胞密度至1.25×106·L-1。取出37℃保温的不同密度的细胞悬液4 mL移入小烧杯中,加入50℃的5%琼脂1 mL,迅速混匀,即配成0.2%琼脂培养基,立即浇入24孔培养板中,每孔加1 mL,置室温使琼脂凝固。培养板移入CO2孵箱,37℃、5%CO2及饱和湿度下培养2~3周。倒置显微镜下计数直径大于75 μm或含50个细胞以上的克隆,并计算形成抑制率。实验重复3次。
1.5 流式细胞仪检测设立空白对照组和药物处理组,药物处理组包括30、60、120 μmol·L-1 DADS分别作用24 h。1 000 r·min-1离心5 min,收集培养细胞,预冷PBS吹打重悬,离心沉淀,重复1次。用4℃预冷的75%乙醇固定细胞,冰盒送检。样本上机前离心洗涤,弃上清,加1 g·L-1碘化丙啶50 μL,振荡混匀,避光置冰箱30 min,上机检测时300目尼龙网滤过,计数10 000个细胞,进行细胞周期和DNA含量分析。实验重复3次。
1.6 Western blot分别收集细胞,冰PBS洗两次,以1×107个细胞浓度加入100 μL裂解液(100 mmol·L-1 NaCl,10 mmol·L-1 Tris-HCl pH 7.6,1 mmol·L-1 EDTA pH8.0,1 mg·L-1 Aprotinin,100 mg·L-1 PMSF),冰上裂解1 h,12 000 r·min-1离心10 min,吸出上清液,即为细胞总蛋白。根据Bradford法,分光光度计在570 nm波长处测定光吸收值,以溶剂为空白对照,牛血清白蛋白为标准绘制曲线,依据标准曲线推算蛋白质含量。收集蛋白,变性后经10%SDS-聚丙烯酰胺凝胶电泳后转移至PVDF膜上,用含5%牛血清白蛋白的TBST(Tris-HCl 20 mmol·L-1,NaCl 137 mmol·L-1含0.1% Tween-20)封闭1h,一抗4℃孵育过夜,TBST洗3次,每次5 min;相应的二抗孵育1 h,TBST洗3次,每次5 min,化学发光剂检测蛋白质印迹,薄层扫描仪测定印迹区带的光密度值。以β-actin作为内参,实验重复3次。
1.7 统计学处理 采用SPSS 15.0统计学软件进行结果处理。实验所得数据采用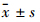 表示,实验组与对照组的区别用方差分析。
表示,实验组与对照组的区别用方差分析。
HL-60细胞群体倍增时间为19.14 h,随着DADS浓度增加到60、120 μmol·L-1,细胞群体倍增时间分别延长到35.03、71.82 h(P<0.05)。Tween 80和30 μmol·L-1 DADS对HL-60细胞群体倍增时间无明显影响 (P>0.05)。
2.2 DADS对HL-60细胞克隆形成的影响Fig1显示,30、60、90、120 μmol·L-1 DADS对HL-60细胞软琼脂克隆形成的抑制率分别为35.06%、62.10%、93.79%、99.35%,抑制作用呈剂量依赖关系 (P<0.05)。Tween 80对HL-60克隆形成无明显影响 (P>0.05)。

|
| Fig 1 Clone formation of HL-60 cells by DADS (×20) A: Control HL-60 cells; B: Tween-80; C: DADS 30 μmol·L-1; D: DADS 60 μmol·L-1; E: DADS 90 μmol·L-1; F: DADS 120 μmol·L-1 |
Fig2显示,60、120 μmol·L-1 DADS作用HL-60细胞24 h后,G2/M期百分率增加到17.6%、22.3%,与对照组比较差异有显著性 (P<0.05)。表明DADS可阻滞HL-60细胞于G2/M期。

|
| Fig 2 Effect of G2/M in HL-60 cells treated by DADS at different time A: Control; B: DADS 30 μmol·L-1; C:DADS 60 μmol·L-1; D:DADS 120 μmol·L-1 |
Fig3显示,60 μmol·L-1 DADS作用HL-60细胞1 h后,与对照组比较,p-Chk1表达呈时间依赖性上调 (P<0.05);Chk1总蛋白表达无改变 (P>0.05)。

|
| Fig 3 Expression of Chk1 and p-Chk1 in HL-60 cells treated by DADS *P<0.05 vs control (0 h) |
Fig4显示,60 μmol·L-1 DADS在作用不同时间后,与对照组比较,Chk2与p-Chk2随时间延长表达无明显差异 (P>0.05)。

|
| Fig 4 Expression of Chk2 and p-Chk2 in HL-60 cells treated by DADS |
Fig5显示,60 μmol·L-1 DADS处理HL-60细胞4 h后,与对照组比较,Cdc25C表达开始下降,到36 h,Cdc25C下调90.16%,DADS呈时间依赖性抑制Cdc25C表达 (P<0.05)。

|
| Fig 5 Effect of DADS on expression of Cdc25C in HL-60 cells *P<0.05 vs control |
Fig6显示,DADS处理4 h后,Cyclin B1的表达即开始下调,DADS处理12 h后,CyclinB1的表达明显下调 (P<0.05);36 h时,与对照组比较,CyclinB1表达明显下降89.02% (P<0.05)。

|
| Fig 6 Effect of DADS on expression of CyclinB1 in HL-60 cells *P<0.05 vs control |
Fig7显示,60 μmol·L-1 DADS呈时间依赖性抑制HL-60细胞CDK1表达 (P<0.05)。DADS处理4 h后,CDK1表达即开始下调,处理8 h后,CDK1表达明显下降,到36 h时,CDK1表达下调34.87% (P<0.05)。

|
| Fig 7 Effect of DADS on expression of CDK1 in HL-60 cells *P<0.05 vs control |
60 μmol·L-1 DADS处理后,随时间的延长,14-3-3蛋白表达较对照组无明显差异 (P>0.05)。
3 讨论本研究采用细胞计数与软琼脂克隆形成实验发现,DADS可明显抑制白血病HL-60细胞增殖,延长细胞群体倍增时间,抑制细胞克隆形成,并随浓度的增加而作用增强。流式细胞术分析显示,60、120 μmol·L-1 DADS作用HL-60细胞24 h后,DADS呈浓度依赖性阻滞HL-60细胞G2/M期,表明DADS抑制HL-60细胞增殖效应与G2/M期阻滞有密切关系。
细胞周期调控在肿瘤的发生和治疗中起着重要的作用。DNA损害有G1/S、S与G2/M检查点,这些检查点能够使DNA容易修复和促进未修复的细胞凋亡。抑制肿瘤细胞检查点,已成为肿瘤治疗的新策略。G2/M期检测点能防止受损的DNA和未完成复制的DNA进入有丝分裂,故G2/M期阻滞显得更为重要。目前,已经研发许多特异性靶向检查点的化合物,其靶点之一是Chk1[7]。目前认为,调控细胞周期G2/M阻滞途径主要是ATR/Chk1/Cdc25C。当DNA发生损伤时,Chk1在丝氨酸317和345位点被ATR磷酸化。活化的Chk1可通过特异性磷酸化Cdc25C,阻止Cdc25C对Cdc2的去磷酸化作用,从而抑制CDK1与Cyclin B复合物的活性,阻滞细胞在G2/M期[8, 9, 10, 11, 12, 13, 14, 15]。Yu 等[8]研究显示,mTOR抑制剂RAD001与辐射结合可通过活化Chk1,抑制CDK1与cyclin B1复合物形成,阻滞口腔癌SCC4细胞于G2/M。NSC746364可通过ATR/Chk1通路下调cyclin B1与激活caspase-3,诱导肺癌A549细胞G2/M阻滞与凋亡[9]。MMEQ可促进Chk1与Cdc25c表达,诱导膀胱癌TSGH8301细胞G2/M阻滞[10]。洋地黄毒苷及其衍生物D6-MA诱导肺癌NCI-H460细胞凋亡与G2/M阻滞和Chk1/2和p53相关蛋白下调有关[11]。鬼臼毒素HY-1可通过ATR-Chk1-Cdc25C通路调控结肠癌HCT116细胞G2/M阻滞[12]。鸡血藤能够通过磷酸化Chk1/Chk2抑制乳腺癌MCF-7细胞和结肠癌HT-29细胞增殖与阻滞在G2/M[13]。Glionitrin A可活化ATM-ATR-Chk1/2通路,上调p-histone 2AX,促进前列腺癌DU145细胞S与G2/M阻滞与凋亡[14]。我们研究显示,DADS可磷酸化Chk1,而不是Chk2,下调Cdc25C与Cyclin B1,阻滞人胃癌BGC823细胞在G2/M期。并且,Chk1转染BGC823细胞可增加G2/M阻滞,而沉默Chk1可消除DADS下调Cdc25C与Cyclin B1和G2/M阻滞。但是,Chk2高表达与沉默Chk2无此作用。表明DADS是通过特异性Chk1磷酸化,而不是Chk2,调控Chk1/Cdc25C/Cyclin B1通路诱导 BGC823细胞G2/M阻滞[15]。
关于白血病的研究,Yuan 等[16]发现,Pim 激酶可在Ser 280磷酸化Chk1诱导急性髓系白血病细胞凋亡与G2/M阻滞。Chk1在热应激诱导白血病Jurkat细胞凋亡与G2/M阻滞起着重要作用,Chk1抑制剂与干扰Chk1可废除热应激诱导白血病Jurkat细胞凋亡与G2/M检查点活化,有趣的是,ATR-Chk1途径优先活化,而不是ATR-Chk2[17]。本研究结果显示,DADS处理HL-60细胞后,p-Chk1表达上调,而p-Chk2没有改变,表明DADS激活Chk1,而不是Chk2,并且,Cdc25C呈时间依赖性表达下降,随即Cyclin B1和CDK1几乎同时下调,表明Chk1活化可抑制Cdc25C活性,影响CDK1与Cyclin B1复合物的活化,从而导致G2/M阻滞,提示DADS通过Chk1/Cdc25C/CyclinB1/CDK1通路导致HL-60细胞阻滞于G2/M期。本研究发现14-3-3蛋白表达没有改变,其是否与Cdc25C结合调控Cdc25C活性,影响Cyclin B1和CDK1作用尚待进一步研究。
| [1] | Jemal A, Bray F, Center M M, et al. Global cancer statistics[J]. CA Cancer J Clin, 2011, 61(2):69-90. |
| [2] | Yi L, Su Q. Molecular mechanisms for the anti-cancer effects of diallyl disulfide[J]. Food Chem Toxicol, 2013, 57:362-70. |
| [3] | Tan H, Ling H, He J, et al. Inhibition of ERK and activation of p38 are involved in diallyl disulfide induced apoptosis of leukemia HL-60 cells[J]. Arch Pharm Res, 2008, 31(6):786-93. |
| [4] | Yi L, Ji X X, Tan H, et al. Involvement of Mcl1 in diallyl disulfide-induced G2/M cell cycle arrest in HL-60 cells. Oncol Rep, 2012, 27(6):1911-7. |
| [5] | Yi L, Ji X X, Lin M, et al. Diallyl disulfide induces apoptosis in human leukemia HL-60 cells through activation of JNK mediated by reactive oxygen[J]. Pharmazie, 2010, 65(9):693-8. |
| [6] | Yi L, Ji X X, Tan H, et al. Role of Ras-related C3 botulinum toxin substrate 2 (Rac2), NADPH oxidase and reactive oxygen species in diallyl disulphide-induced apoptosis of human leukaemia HL-60 cells[J]. Clin Exp Pharmacol Physiol, 2010, 37(12):1147-53. |
| [7] | Bucher N, Britten C D. G2 checkpoint abrogation and checkpoint kinase-1 targeting in the treatment of cancer[J]. Br J Cancer, 2008, 98(3):523-8. |
| [8] | Yu C C, Hung S K, Liao H F, et al. RAD001 enhances the radiosensitivity of SCC4 oral cancer cells by inducing cell cycle arrest at the G2/M checkpoint[J]. Anticancer Res, 2014, 34(6):2927-35. |
| [9] | Chung Y L, Pan C H, Liou W H, et al. NSC746364, a G-quadruplex-stabilizing agent, suppresses cell growth of A549 human lung cancer cells through activation of the ATR/Chk1-dependent pathway[J]. J Pharmacol Sci, 2014, 124(1):7-17. |
| [10] | Hsu S C, Yu C C, Yang J S, et al. A novel synthetic 2-(3-methoxyphenyl)-6, 7-methylenedioxoquinolin-4-one arrests the G2/M phase arrest via Cdc25c and induces apoptosis through caspase-and mitochondria-dependent pathways in TSGH8301 human bladder cancer cells[J]. Int J Oncol, 2012, 40(3):731-8. |
| [11] | Elbaz H A, Stueckle T A, Wang H Y, et al. Digitoxin and a synthetic monosaccharide analog inhibit cell viability in lung cancer cells[J]. Toxicol Appl Pharmacol, 2012, 258(1):51-60. |
| [12] | Zhao Y, Wu Z, Zhang Y, et al. HY-1 induces G(2)/M cell cycle arrest in human colon cancer cells through the ATR-Chk1-Cdc25C and Weel pathways[J]. Cancer Sci, 2013, 104(8):1062-6. |
| [13] | Wang Z Y, Wang D M, Loo T Y, et al. Spatholobus suberectus inhibits cancer cell growth by inducing apoptosis and arresting cell cycle at G2/M checkpoint[J]. J Ethnopharmacol, 2011, 133(2):751-8. |
| [14] | Kim Y J, Park H B, Yoo J H, et al. Glionitrin A, a new diketopiperazine disulfide, activates ATM-ATR-Chk1/2 via 53BP1 phosphorylation in DU145 cells and shows antitumor effect in xenograft model[J]. Biol Pharm Bull, 2014, 37(3):378-86. |
| [15] | Bo S, Hui H, Li W, et al. Chk1, but not Chk2, is responsible for G2/M phase arrest induced by diallyl disulfide in human gastric cancer BGC823 cells[J]. Food Chem Toxico, 2014, 68:61-70. |
| [16] | Yuan L L, Green A S, Bertoli S, et al. Pim kinases phosphorylate Chk1 and regulate its functions in acute myeloid leukemia[J]. Leukemia, 2014, 28(2):293-301. |
| [17] | Furusawa Y, Iizumi T, Fujiwara Y, et al. Inhibition of checkpoint kinase 1 abrogates G2/M checkpoint activation and promotes apoptosis under heat stress[J]. Apoptosis, 2012, 17(1):102-12. |