



2. 中山大学附属第三医院放射科, 广东 广州 510630;
3. 中山大学附属第一医院麻醉科, 广东 广州 510089
2. Dept of Radiology, the Third Affiliated Hospital, Sun Yet-sen University, Guangzhou 510630, China;
3. Dept of Anesthesiology, the First Affiliated Hospital, Sun Yet-sen University, Guangzhou 510089, China
偏移的CD8+T细胞极化在许多免疫异常反应中起重要作用,比如风湿性关节炎[1]、糖尿病[2]和颅内感染[3]。在哮喘发生中,CD8+T细胞极化同样起重要作用,但何种原因导致其极化并不清楚[4]。
胰岛素样生长因子(insulin-like growth factor-1,IGF1)又称为生长调节素C(somatomedin C),促进生长和防止细胞死亡。IGF1在人的一生中都发挥作用,尤其是在儿童期和青春期。IGF1主要由肝脏产生,通常是生长激素启动的后续信号。IGF1也是一些过敏性疾病(如哮喘)的发病因素之一。嗜碱性粒细胞表达IGF1受体,当暴露于IGF1后,前者被激活并释放胞内颗粒[5]。小鼠气道过敏模型中检出IGF1高表达[6],还有学者认为IGF1对未成熟的肺成纤维细胞有促进α平滑肌表达和胶原蛋白合成的作用,这将导致肺纤维化(由核因子κB转录介导)[7]。除肝脏外,IGF1还可由气道炎症细胞产生,但具体是何种类型细胞尚不清楚[8]。
被激活后T细胞通过凋亡进入激活诱导的细胞死亡(activation-induced cell death,AICD),维持机体免疫平衡。作为死亡信号受体的Fas基因及其配体FasL,在T细胞被激活后表达上调,Fas/FasL相互作用下启动细胞凋亡[9]。除了Fas/FasL通路外,还有其他分子在凋亡启动中发挥作用,如p53蛋白。尽管p53蛋白主要是诱导肿瘤细胞凋亡,其他细胞中也有表达,氮芥诱导的小鼠皮肤细胞凋亡和炎症反应,这一过程就是依赖p53基因表达的[10]。紫外线照射诱导人角质细胞调节ERK1/2和血栓调节素表达就是由p53基因介导的[11]。然而,p53基因在CD8+T细胞AICD过程,尤其是偏移CD8+T细胞极化中的作用尚不清楚。我们发现,过敏性刺激使小鼠气道上皮细胞表达IGF1,这种上皮细胞产生的IGF1抑制CD8+T细胞p53表达,参与抗原特异性的CD8+T细胞极化过程。
1 材料与方法 1.1 试剂IGF1及p53抗体购自Santa Cruz Biotech上海分公司;定量qRT-PCR及Western免疫印迹试剂购自Invitrogen上海分公司;Der p1蛋白购自MyBiosource公司(San Diego,CA);颗粒酶B及穿孔素ELISA套件购自R&D Systems上海分公司;膜联蛋白Annexin V套件购自Sigma Aldrich上海分公司;免疫细胞分离提取套件购自Miltenyi Biotech上海分公司;DNA纯化套件购自Promega北京分公司;EpiXploreTM甲基化检测套件购自Clontech北京分公司。
1.2 细胞培养人鼻腔上皮细胞株RPMI2650购自美国细胞培养协会(American Type Culture Collection,ATCC,Manassas,VA,USA),置于DMEM细胞培养基(Dulbecco′s Modified Eagle Medium)培养,加入10%小牛血清、100 kU·L-1青霉素、100 mg·L-1链霉素及200 mmol·L-1谷氨酸。培养箱条件为37℃恒温,含5% CO2。
1.3 定量实时PCR(qRT-PCR)细胞总RNA通过TRIzol reagents试剂盒提取。cDNA由逆转录试剂盒合成。qRT-PCR由MiniOpticon实时PCR系统(Bio-Rad Life Science公司,上海)完成。研究所用引物为:IGF1 (NCBI: NM_001111283),正向:3′-TCTGAATCTTGGCTGCTGGA-5′,反向: 3′-TGTGCTTCTTGACGACTTGC-5′;p53 (NCBI: AB082923),正向:3′-TGGCCATCTACAAGCAGTCA-5′,反向: 3′-GGTACAGTCAGAGCCAACCT-5′。结果用内参β-actin相对量表达。
1.4 Western免疫印迹细胞总蛋白提取后由SDS-PAGE凝胶电泳分离,再转印至硝酸纤维膜上。经5%脱脂牛奶孵育封闭后加入一抗(100~500 μg·L-1),室温下孵育1 h后,加入辣根过氧化物酶结合二抗继续孵育1 h。每次孵育完成后,用缓冲液(Triton X-100)冲洗多余抗体。免疫反应结束后生成化学发光底物,并由X射线胶片成像。胶片成像后由系统自带图像软件量化免疫印迹密度。
1.5 流式细胞计数细胞经荧光标记的一抗染色后由流式细胞计数仪(FACSCanto II,BD Bioscience,上海分公司)计数。凋亡细胞通过Annexin V套件染色后检测。死亡细胞由碘化丙啶(propidium iodide,PI)5 mg·L-1,15 min染色后检测计数。
1.6 CD8+T细胞纯化由健康志愿者获得外周血标本(每人20 mL),实验及血样采集经我院伦理委员会批准。经梯度离心获得外周血单核细胞(peripheral blood mononuclear cells,PBMC)。通过免疫磁珠细胞分离套件将PBMC中CD8+T细胞及CD11c+树突状细胞(dendritic cells,DC)分离,并经流式细胞仪检测,CD8+CD25-T细胞及DC纯度均超过98%。
1.7 抗原特异性CD8+T细胞多克隆增殖的评估将CD8+CD25-T细胞用琥珀酰亚胺酯(carboxy fluorescein succinimidyl ester,CFSE)标记,置于2 mg·L-1 CD3抗体和 5 mg·L-1 CD28抗体包被环境下培养72 h。T细胞增殖率通过CFSE稀释法测定。
1.8 抗原特异性CD8+T细胞激活的评估将CD8+CD25-T细胞置于Transwell底层小室中培养,间隔层为RPMI2650细胞株。Der p1蛋白(盐水作为对照或加入抗IGF1中和抗体200 μg·L-1)置于顶层小室,浓度为30 μg·L-1,孵育72 h。收获CD8+T细胞并用CFSE标记,加入DC细胞(T细胞 ∶ DC=5 ∶ 1),在30 μg·L-1 Der p1蛋白或小牛血清蛋白中培养72 h。CD8+T细胞增殖率通过CFSE稀释法测定。培养液中颗粒酶B及穿孔素水平由相应ELISA试剂盒检测。
1.9 51铬(51Cr)释放检测激活的CD8+T细胞用51Cr释放检测法分析。以结肠癌细胞株T84作为靶细胞,用51Cr标记孵育2 h。冲洗后将T84细胞(108·L-1)和CD8+T细胞(108·L-1)置于Transwell底层小室中。间隔层为RPMI2650细胞(109·L-1)。底层小室上清液每4 h收集一次75 μL。51Cr活性由γ射线计数器(Perkin Elmer公司,美国)测定,最大释放量和自发释放量分别通过加入2% Triton X-100和培养液获得。CD8+T细胞激活产生的51Cr特异性释放量由以下公式计算:(A-B)/(C-B)×100%,A:实际测定量,B:自发释放量,C:最大释放量。
1.10 p53基因甲基化测定将纯化的DNA用EpiXploreTM甲基化检测套件进行亚硫酸盐转化后,进行甲基化测定。p53基因的引物序列分别为:(1)甲基化引物,正向:3'-ATTTTAGTGGTAATTTATTGGGACG-5',反向: 3'-TAATAAAACTCCCCTTTCTTACGAA-5';(2)非甲基化引物,正向:3'-TTTTAGTGGTAATTTATTGGGATGG-5',反向: 3'-TAATAAAACTCCCCTTTCTTACAAA-5'。
1.11 统计学分析文中所涉及细胞实验数据,如无特殊说明,均代表3次独立实验结果。数据以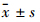 形式呈现,两组间差异比较用t检验,两组以上组间差异比较用单因素方差分析。
形式呈现,两组间差异比较用t检验,两组以上组间差异比较用单因素方差分析。
气道上皮细胞能够产生IGF1[12],但其触发生成的诱因尚未明确。我们同时检测了IGF1在人气道上皮细胞(RPMI2650细胞)基线水平的mRNA(Fig1A)和蛋白(Fig1B-C)水平的表达量,二者在暴露于Der p1后均明显增加,并且这种增加具有剂量依赖性(Fig1)。该结果提示:鼠致敏原能上调人气道上皮细胞IGF1表达。

|
| Fig 1 Airway epithelial cells express IGF1 Human airway epithelial cell line RPMI2650 cells, was cultured in the presence or absence of a mite allergen, Der p1, for 72 h. A: The bars indicate the levels of IGF1 mRNA; B: The immune blots indicate the IGF1 protein levels. The bars below the blots indicate the summarized integrated density of the blots. BSA is an irrelevant control protein (50 μg·L-1); C: The bars indicate the cell counts from day 0 to day 3. **P<0.01 vs BSA group (A, B), or day 0 group (C). |
尽管偏移CD8+T细胞极化在气道过敏性疾 病发生中起作用[13],但上皮细胞表达的IGF1是否对CD8+T细胞极化有作用尚不清楚。我们发现,原始CD8+T细胞能够被CD3/CD28抗体激活,表现为CD8+T细胞增殖(Fig2A),同时激活的CD8+T细胞中存在大量凋亡细胞(Fig2B-C)。在混合培养液中,盐水对照组中凋亡CD8+T细胞处于低水平(Fig2B、2Ca),加入CD3/CD28抗体激活后凋亡CD8+T细胞明显增加(Fig2B、2Cb)。加入Der p1孵育的上皮细胞后,凋亡细胞增加的趋势被抑制(Fig2B、2Cc),而加入未经处理的上皮细胞则无此效果(Fig2B、2Cd)。该结果提示,气道上皮细胞生成的IGF1会抑制CD8+T细胞AICD过程。为进一步证实该想法,我们在上述实验中加入IGF1抗体,发现Der p1孵育上皮细胞对CD8+T细胞凋亡抑制作用被阻断了(Fig2B、2Ce、2Cf),加入重组IGF1进一步证实了该结果(Fig2B、2Cg)。

|
| Fig 2 Epithelial cell-derived IGF1 reduces AICD of CD8+T cells A: The histograms indicate the proliferation of CD8+T cells; B: The bars indicate the frequency of dead CD8+T cells and apoptotic CD8+T cells in the activated CD8+T cells; C: The dot plots indicate the frequencies of PI+ or/and Annexin V+ CD8+T cells. **P<0.01 vs group a. |
p53基因产物能够诱导细胞凋亡,由此我们进一步确定IGF1是否通过该途径抑制CD8+T细胞AICD[14]。我们利用实验Fig2中部分细胞,检测其生成的IGF1能否调节CD8+T细胞p53蛋白表达。定量PCR和Western免疫印迹均显示CD8+T细胞p53蛋白表达。以CD3/CD28抗体激活后,CD8+T细胞p53蛋白表达明显增加,Der p1孵育的RPMI2650细胞能抑制该趋势,而加入IGF1抗体后这种作用消失(Fig3A-B)。由此可见,激活CD8+T细胞将使p53蛋白表达增加,而上皮细胞生成的IGF1则可以阻断该过程。
为阐明IGF1抑制CD8+T细胞p53表达的机制,我们进行了甲基化特异性PCR检测。结果显示,在原始CD8+T细胞中,p53基因存在部分甲基化,而在暴露于重组IGF1后,这一比例明显增加。在MEK-ERK缺乏CD8+T细胞中则未观察到该现象(Fig3C)。Western免疫印迹显示,IGF1处理后的CD8+T细胞Bcl-2和Bcl-xL蛋白水平并无明显变化(Fig3D-E)。

|
| Fig 3 IGF1 enhances methylation of p53 gene Nave CD8+T cells were treated in the same procedures of Fig 2C, the bars (A) and immune blots (B) indicate the levels of p53 in CD8+T cells. **P<0.01 vs group a. C:The p53 gene methylation results. M: Methylated; U: Un-methylated; rIGF1: Recombinant IGF1; U0126: The MEK-ERK inhibitor; LY294002:The PIK3-Akt inhibitor. BSA is an irrelevant protein using as a control; Anti-CD3/CD28: CD8+T cells were treated with anti-CD3/CD28 antibodies. D and E: The immune blots indicate the protein contents of Bcl-2 (D) and Bcl-xL (E) in the CD8+T cell extracts. |
气道上皮细胞表达MHC I和MHC II[15]及B7分子,并将抗原呈递给T细胞[14]。 为了明确气道上皮细胞生成的IGF1能否作为抗原特异性偏移CD8+T细胞极化的启动因子,我们将原始CD8+T细胞和Der p1蛋白诱导的气道上皮细胞共同孵育48 h。将获得的CD8+T细胞置于特异性抗原Der p1环境下,加入新鲜提取的DC共孵育72 h。特异性CD8+T细胞出现明显 增殖,加入IGF1中和抗体后增殖被抑制(Fig4A)。培养液中细胞毒因子、颗粒酶B(Fig4B)及穿孔素(Fig4C)水平明显升高。该结果提示,气道上皮细胞生成的IGF1能够触发抗原特异性CD8+T细胞极化。CD8+T细胞的激活通过特异性溶解反应得到证实(Fig4D)。

|
| Fig 4 Epithelial cell-derived IGF1 induces CD8+T cell polarization A~F:The histograms indicate the frequency of CD8+T cell proliferation; A~B: Without the presence of epithelial cells; C~E: In the presence of epithelial cells; F: Staining controls; G~I: The bars indicate the levels of granzyme B (G), perforin (H) and specific lysis (I). The labels of x axis are the same as the labels of histograms. **P<0.01 vs group A. |
许多免疫系统异常或多或少都与偏移CD8+T 细胞极化相关。本研究显示气道上皮细胞(RPMI 2650细胞),能够在鼠过敏原Der p1蛋白作用下产生IGF1。而这种气道上皮细胞生成的IGF1将使p53基因发生甲基化,进而抑制CD8+T细胞的凋亡进程,促成CD8+T细胞极化。
鼠过敏原在许多过敏性疾病发生过程中扮演重要角色。除了作为通常意义下的过敏原外,它们中的一些亚型(如Der p1)具有蛋白酶样作用,能够影响上皮细胞紧密连接处的完整性,并且能够激活蛋白酶活化受体2,从而破坏上皮屏障功能[16]。我们的研究提供了进一步证据:Der p1能够诱导RPMI2650细胞生成IGF1。IGF1通常是由肝细胞生成的,但其他细胞(如气道组织中的炎症细胞)也可以产生[8]。
AICD是一种T细胞的正常激活反应。这种过程在T细胞被激活后就启动并诱导凋亡,AICD在维持机体免疫系统稳定中意义重大。AICD是由Fas受体(Fas,CD95)和Fas配体(FasL,CD95 ligand)相互作用下诱导的细胞凋亡。机体许多(免疫耐受)重要器官,如睾丸、大脑、角膜等,其细胞均有FasL高表达,用以抑制免疫反应对这些器官可能造成的损伤。这就不难理解如果AICD机制出现异常,会发生多种自身免疫疾病。本研究显示,被CD3/CD28抗体激活后凋亡CD8+T细胞明显增加,而气道上皮细胞生成的IGF1使凋亡和死亡细胞数减少。加入IGF1中和抗体后,减少CD8+T细胞凋亡的作用被抑制,再加入重组IGF1又可恢复这种抑制作用。其他学者也发现IGF1能够抑制细胞凋亡[17, 18]。
p53基因作用缺失会造成许多疾病。p53蛋白是一种肿瘤抑制蛋白,在许多肿瘤中存在变异[19]。p53蛋白还在其他疾病中起作用,诸如缺血性细胞损伤[20]、吸烟相关性疾病[21]。我们发现p53蛋白在气道上皮细胞生成的IGF1诱导抑制CD8+T细胞AICD过程中有作用。IGF1能够通过MEK-ERK通路诱发p53基因甲基化。其他学者证实在间质干细胞向成骨细胞转化时,IGF1同样通过MEK-ERK通路促进这一进程[22]。
总之,鼠过敏原Der p1蛋白能够诱发气道上皮细胞——RPMI2650细胞产生IGF1,后者通过诱导p53基因甲基化抑制CD8+T细胞AICD。
| [1] | Carvalheiro H, da Silva J A, Souto-Carneiro M M. Potential roles for CD8(+) T cells in rheumatoid arthritis[J]. Autoimmun Rev, 2013, 12(3):401-9. |
| [2] | Coppieters K T, von Herrath M G. Viruses and cytotoxic T lymphocytes in type 1 diabetes[J]. Clin Rev Allergy Immunol, 2011, 41(2):169-78. |
| [3] | Willing A, Friese M A. CD8-mediated inflammatory central nervous system disorders[J]. Curr Opin Neurol, 2012, 25(3):316-21. |
| [4] | Robinson D S. The role of the T cell in asthma[J]. J Allergy Clin Immunol, 2010, 126(6):1081-91. |
| [5] | Koketsu R, Suzukawa M, Kawakami A, et al. Activation of basophils by stem cell factor:comparison with insulin-like growth factor-I[J]. J Investig Allergol Clin Immunol, 2008, 18(4):293-9. |
| [6] | Jaradat M, Stapleton C, Tilley S L, et al. Modulatory role for retinoid-related orphan receptor alpha in allergen-induced lung inflammation[J]. Am J Respir Crit Care Med, 2006, 174(12):1299-309. |
| [7] | Chetty A, Cao G J, Nielsen H C. Insulin-like growth factor-I signaling mechanisms, type I collagen and alpha smooth muscle actin in human fetal lung fibroblasts[J]. Pediatr Res, 2006, 60(4):389-94. |
| [8] | Vieira R P, Duarte A C, Claudino R C, et al. Creatine supplementation exacerbates allergic lung inflammation and airway remodeling in mice[J]. Am J Respir Cell Mol Biol, 2007, 37(6):660-7. |
| [9] | Lettau M, Paulsen M, Schmidt H, et al. Insights into the molecular regulation of FasL (CD178) biology[J]. Eur J Cell Biol, 2011, 90(6-7):456-66. |
| [10] | Inturi S, Tewari-Singh N, Jain A K, et al. Absence of a p53 allele delays nitrogen mustard-induced early apoptosis and inflammation of murine skin[J]. Toxicology, 2013, 311(3):184-90. |
| [11] | Huang H C, Chang T M, Chang Y J, et al. UVB irradiation regulates ERK1/2-and p53-dependent thrombomodulin expression in human keratinocytes[J]. PLoS One, 2013, 8(7):e67632. |
| [12] | Kawaguchi M, Fujita J, Kokubu F, et al. Induction of insulin-like growth factor-I by interleukin-17F in bronchial epithelial cells[J]. Clin Exp Allergy, 2010, 40(7):1036-43. |
| [13] | Dakhama A, Collins M L, Ohnishi H, et al. IL-13-producing BLT1-positive CD8 cells are increased in asthma and are associated with airway obstruction[J]. Allergy, 2013, 68(5):666-73. |
| [14] | Pant V, Quintas-Cardama A, Lozano G. The p53 pathway in hematopoiesis:lessons from mouse models, implications for humans[J]. Blood, 2012, 120(26):5118-27. |
| [15] | Kreisel D, Lai J, Richardson S B, et al. Polarized alloantigen presentation by airway epithelial cells contributes to direct CD8+T cell activation in the airway[J]. Am J Respir Cell Mol Biol, 2011, 44(6):749-54. |
| [16] | Roelandt T, Heughebaert C, Hachem J P. Proteolytically active allergens cause barrier breakdown[J]. J Invest Dermatol, 2008, 128(8):1878-80. |
| [17] | Chung H, Seo S, Moon M, et al. IGF-I inhibition of apoptosis is associated with decreased expression of prostate apoptosis response-4[J]. J Endocrinol, 2007, 194(1):77-85. |
| [18] | Garrouste F, Remacle-Bonnet M, Fauriat C, et al. Prevention of cytokine-induced apoptosis by insulin-like growth factor-I is independent of cell adhesion molecules in HT29-D4 colon carcinoma cells-evidence for a NF-kappaB-dependent survival mechanism[J]. Cell Death Differ, 2002, 9(7):768-79. |
| [19] | Muller P A, Vousden K H. p53 mutations in cancer[J]. Nat Cell Biol, 2013, 15(1):2-8. |
| [20] | Hong L Z, Zhao X Y, Zhang H L. p53-mediated neuronal cell death in ischemic brain injury[J]. Neurosci Bull, 2010, 26(3):232-40. |
| [21] | Yu M W, Yang S Y, Chiu Y H, et al. A p53 genetic polymorphism as a modulator of hepatocellular carcinoma risk in relation to chronic liver disease, familial tendency, and cigarette smoking in hepatitis B carriers[J]. Hepatology, 1999, 29(3):697-702. |
| [22] | Xue P, Wu X, Zhou L, et al. IGF1 promotes osteogenic differentiation of mesenchymal stem cells derived from rat bone marrow by increasing TAZ expression[J]. Biochem Biophys Res Commun, 2013, 433(2):226-31. |