 2021, Vol. 32
2021, Vol. 32扩展功能
文章信息
- 王卓, 吴琼, 金美松, 邢悦鹏, 杨义军, 吴益民
- WANG Zhuo, WU Qiong, JIN Mei-song, XING Yue-peng, YANG Yi-jun, WU Yi-min
- 中国长白山地区啮齿动物巴尔通体感染调查及基因型分析
- An investigation of Bartonella infection in rodents in the Changbai Mountains in China and its genotype analysis
- 中国媒介生物学及控制杂志, 2021, 32(5): 594-598
- Chin J Vector Biol & Control, 2021, 32(5): 594-598
- 10.11853/j.issn.1003.8280.2021.05.017
-
文章历史
- 收稿日期: 2021-03-28




2 长春市城乡规划研究院环境检测中心, 吉林 长春 130021;
3 辽宁省食品检验检测院, 辽宁 沈阳 110016
2 Centre of Environmental Detection, Changchun Academy of Plan and Design for Urban and Rural, Changchun, Jilin 130021, China;
3 Liaoning Institute for Food Control, Shenyang, Liaoning 110016, China
巴尔通体属于变形菌纲a亚群、根瘤菌目、巴尔通体科、巴尔通体属(Bartonella)。目前,巴尔通体属已发现30多种和3亚种及一些未命名的巴尔通体。目前认为与人类疾病有关的巴尔通体至少有12种和2亚种,其中有6种〔阿尔萨斯巴尔通体(B. alsatica)、伊丽莎白巴尔通体(B. elizabethae)、格拉汉姆巴尔通体(B. grahamii)、塔麦巴尔通体(B. tamiae)、文森巴尔通体阿鲁潘亚种(B. vinsonii subsp.arupensis)和瓦氏巴尔通体(B. washoensis)〕与小型哺乳动物,特别是野生啮齿动物密切相关[1],可引起人类心内膜炎、猫抓病、杆状样血管瘤及脑膜炎等[1-4]。
巴尔通体感染属于人兽共患的自然疫源性疾病。研究表明,巴尔通体储存宿主广泛,啮齿类动物是重要的储存宿主[5]。我国自2000年首次证实云南省鼠群中巴尔通体流行[6],继而广东省、内蒙古自治区等多地报道从啮齿类动物分离或检测到巴尔通体[7-12],表明南方和北方地区宿主动物巴尔通体感染普遍。东北地区地域辽阔,地理环境复杂,生物资源丰富,是我国重要的鼠源传染病自然疫源地。了解与掌握啮齿动物巴尔通体自然感染状况和特点,可为该病的预防控制及疫源地证实提供科学依据。本研究在我国东北长白山地区选点采集啮齿动物样本,应用PCR方法进行巴尔通体感染调查与基因型特征分析。
1 材料与方法 1.1 鼠标本采集选择长白山脉北端黑龙江省密山县(2016年,耕地),长白山脉中、南端吉林省集安市,辽宁省宽甸县(2012年,林区和林缘耕地),采用夹夜法和笼夜法捕获啮齿动物。现场分类鉴定,消毒处理后解剖取脾和肝脏,置于细胞冻存管内,液氮保存。运回实验室后-80 ℃保存。
1.2 鼠脾标本DNA提取鼠脾经玻璃研磨器中充分研碎,采用DNeasy Blood & Tissue Kit(QIAGEN)按产品说明书操作提取DNA,DNA提取液置-20 ℃保存。
1.3 PCR扩增用PCR扩增巴尔通体rpoB基因(RNA聚合酶亚单位基因),rpoB基因引物的设计和扩增条件按文献[13]进行,引物由生工生物工程(上海)股份有限公司合成。PCR rpoB基因正向引物Bar-rpoF:5'-GCACGATTYGCATCATCATTTTCC-3',反向引物Bar-rpoR:5'-CGCATTATGGTCGTATTTGTCC-3',预期扩增目的片段333 bp。扩增产物经1.2%琼脂糖凝胶电泳,Dolphin-Chemi凝胶成像系统分析,照相。
1.4 序列比对及数据分析为进一步鉴定所检测到的巴尔通体基因序列,将所有检出PCR阳性产物送至生工生物工程(上海)股份有限公司进行测序,将所测得的序列结果在美国国立生物技术信息中心(NCBI)利用核酸比对程序BLAST与GenBank中注册的巴尔通体rpoB基因序列进行同源性分析,然后利用MEGA 7.0软件构建系统进化树。
1.5 统计学分析采用SPSS 19.0软件对数据进行统计学分析,率的比较采用χ2检验,P < 0.05为差异有统计学意义。
2 结果 2.1 啮齿动物种类与分布在长白山地区3个调查点共捕获啮齿动物5种178只,其中黑线姬鼠(Apodemus agrarius)126只,占捕获总数的70.79%;大林姬鼠(A. peninsulae)、大仓鼠(Tscherskia triton)和褐家鼠(Rattus norvegicus)分别占13.48%(24/178)、8.43%(15/178)和6.74%(12/178);小林姬鼠(A. sylvaticua)占0.56%(1/178)。
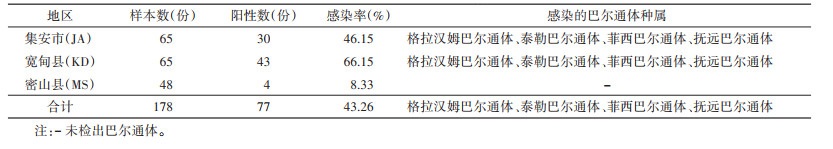
2.2 啮齿动物感染巴尔通体调查3个调查点共检测鼠脾标本178份,PCR检测巴尔通体阳性77份,感染率为43.26%(表 1)。其中,宽甸县野鼠阳性率最高为66.15%,密山县最低为8.33%,各地区野鼠感染率差异有统计学意义(χ2=37.957,P < 0.001)。从表 1可见,集安市和宽甸县野鼠都存在格拉汉姆巴尔通体、泰勒巴尔通体(B. taylorii)、菲西巴尔通体(B. phoceensis)、抚远巴尔通体(B. fuyuanensis)等4种巴尔通体。密山县4份阳性样本测序未成功。
|
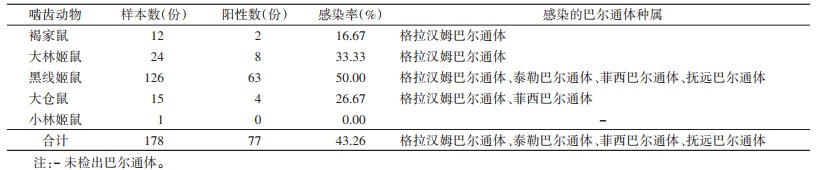
5个鼠种(178只野鼠样本)中,巴尔通体检出阳性率为43.26%,褐家鼠、大林姬鼠、黑线姬鼠和大仓鼠巴尔通体感染率分别为16.67%、33.33%、50.00%和26.67%,小林姬鼠未检出阳性(表 2),不同鼠种间感染率差异有统计学意义(χ2=8.420,P=0.038)。由表 2可见,从不同鼠种中检测到4种巴尔通体种感染,大林姬鼠、黑线姬鼠、褐家鼠和大仓鼠都检测到格拉汉姆巴尔通体,从黑线姬鼠和大仓鼠发现菲西巴尔通体,而抚远巴尔通体和泰勒巴尔通体仅从黑线姬鼠检出。
|
PCR检测野鼠巴尔通体阳性样本77份,成功测序48份,与GenBank数据库中注册的巴尔通体rpoB基因序列进行同源性比较并构建遗传进化树(图 1)。基于rpoB基因共鉴定出4种巴尔通体,分别为格拉汉姆巴尔通体、泰勒巴尔通体、菲西巴尔通体和抚远巴尔通体。以JA76、KD60、JA73、KD3、JA54为代表的27份样本与格拉汉姆巴尔通体及其韩国株相互聚集,同处一分支中,同源性为99.15%~99.71%;但KD58同源性为96.26%。样本JA63、KD21与抚远巴尔通体处同一分支,同源性为98.85%。以KD50、JA64为代表的4份样本与菲西巴尔通体处同一分支,同源性在98.50%以上。以KD57、KD52、JA50为代表的12份样本与泰勒巴尔通体相互聚集在同一分支中,同源性为96.80%~99.42%。另外,样本JA20和JA15未与已知巴尔通体相互聚集,经BLAST验证与格拉汉姆巴尔通体同源性为97.00%。

|
| 图 1 基于巴尔通体rpoB基因(333 bp)构建的分子遗传进化树 Figure 1 The phylogenetic tree based on the rpoB gene (333 bp) of Bartonella |
| |
巴尔通体以吸血节肢动物(蚤、白蛉和蜱等)为传播媒介,宿主动物广泛,野生啮齿动物是巴尔通体的重要自然宿主,呈全球性分布[1, 5]。研究证实,在亚洲的日本、韩国、泰国及我国台湾地区的啮齿动物中巴尔通体感染高度流行(28.80%~56.30%)[5],俄罗斯西伯利亚地区鼠类感染率高达67.00%[14]。近年来国内研究表明,福建和广东省鼠形动物巴尔通体感染率分别为12.34%和19.47%,检出3~6种巴尔通体基因型[8-9];宁夏回族自治区啮齿动物巴尔通体感染率为21.79%,以阿拉善黄鼠(Spermophilus alaschanicus)感染率最高(45.45%)[10];内蒙古地区感染率为38.37%,以达乌尔黄鼠(S. dauricus)阳性率最高(75.00%)[11],北方地区感染率似高于南方部分地区,证明我国许多省(自治区)鼠群巴尔通体感染普遍。地区不同、宿主动物种群不同,巴尔通体感染率及基因型分布也各不相同[6-12]。
本次研究应用PCR方法调查长白山脉3个调查点啮齿动物巴尔通体流行情况,结果表明巴尔通体感染率(43.26%)明显高于南方部分地区,与云南省(44.30%)[6]、黑龙江省黑瞎子岛(57.70%)[12]的报道相近,说明长白山地区鼠群巴尔通体感染高度流行。通过序列测定与遗传进化分析从4个鼠种共检出4种巴尔通体基因型,显示出长白山地区存在巴尔通体基因型别的多样性和宿主动物的多样性特点。调查研究表明该地区啮齿动物巴尔通体感染以格拉汉姆巴尔通体为主(62.50%),感染率以黑线姬鼠最高(50.00%)。Li等[12]从中俄边境黑瞎子岛黑线姬鼠分离到抚远巴尔通体,经鉴定与已知的巴尔通体基因型不同,认为是一新基因型,本研究亦从长白山地区黑线姬鼠检测到抚远巴尔通体,提示抚远巴尔通体可能分布于整个东北地区,而其宿主动物是否存在相对特异性及致病性需进一步调查研究。
本研究采用巴尔通体遗传进化分析常用的rpoB基因,但仅应用单基因分析易影响巴尔通体检测和型别的判断。因此,进一步选用多个基因或多位点序列分析可有效提高检测的准确性和种属间的差异性。
根据本次调查研究结果,长白山地区啮齿动物巴尔通体感染高度流行,存在4种巴尔通体基因型,以格拉汉姆巴尔通体感染为主,巴尔通体感染率以黑线姬鼠最高,并显示出巴尔通体基因型别的多样性和宿主动物的多样性特点。已经证实格拉汉姆巴尔通体是人类巴尔通体感染的病原体,野生啮齿动物可为格拉汉姆巴尔通体的贮存宿主,该基因型在长白山地区分布普遍。随着旅游业的发展和户外活动增加,人类与啮齿动物接触及被蜱、蚤等吸血节肢动物叮咬的机会增加,巴尔通体感染概率将会增加,因此对该疾病的预防与治疗应引起高度重视。
利益冲突 无
| [1] |
Kim KS, Inoue K, Kabeya H, et al. Prevalence and diversity of Bartonella species in wild small mammals in Asia[J]. J Wildlife Dis, 2016, 52(1): 10-21. DOI:10.7589/2015-01-015 |
| [2] |
Kosoy M, Morway C, Sheff KW. Bartonella tamiae sp. nov., a newly recognized pathogen isolated from three human patients from Thailand[J]. J Clin Microbiol, 2008, 46(2): 772-775. DOI:10.1128/JCM.02120-07 |
| [3] |
Fenollar F, Sire S, Raoult D. Bartonella vinsonii subsp. arupensis as an agent of blood culture-negative endocarditis in a human[J]. J Clin Microbiol, 2005, 43(2): 945-947. DOI:10.1128/JCM.43.2.945-947.2005 |
| [4] |
Probert W, Louie JK, Tucker JR, et al. Meningitis due to a "Bartonella washoensis"-like human pathogen[J]. J Clin Microbiol, 2009, 47(7): 2332-2335. DOI:10.1016/j.actatropica.2010.02.015 |
| [5] |
Jiyipong T, Jittapalapong S, Morand S, et al. Bartonella species in small mammals and their potential vectors in Asia[J]. Asian Pac J Trop Biomed, 2014, 4(10): 757-767. DOI:10.12980/APJTB.4.2014C742 |
| [6] |
白瑛, KosoyMY, MaupinGO, 等. 首次证实巴尔通体在我国云南鼠群中流行[J]. 中国人兽共患病杂志, 2002, 18(3): 5-9. Bai Y, Kosoy MY, Maupin GO, et al. Discovery of Bartonella species in rodents in Yunnan[J]. Chin J Zoonoses, 2002, 18(3): 5-9. DOI:10.3969/j.issn.1002-2694.2002.03.001 |
| [7] |
凌锋, 丁丰, 龚震宇, 等. 浙江省啮齿动物巴尔通体感染状况调查[J]. 中国媒介生物学及控制杂志, 2014, 25(1): 24-27. Ling F, Ding F, Gong ZY, et al. Investigation of Bartonella infection in rodents in Zhejiang province, China[J]. Chin J Vector Biol Control, 2014, 25(1): 24-27. DOI:10.11853/j.issn.1003.4692.2014.01.007 |
| [8] |
肖方震, 林代毕, 周淑姮, 等. 福建省鼠类感染巴尔通体调查及序列分析[J]. 中国人兽共患病学报, 2017, 33(7): 607-612. Xiao FZ, Lin DB, Zhou SH, et al. Investigation and sequence analysis on Bartonella infection in rodents in Fujian province, China[J]. Chin J Zoonoses, 2017, 33(7): 607-612. DOI:10.3969/j.issn.1002-2694.2017.07.007 |
| [9] |
李雪云, 陈经雕, 黎聪荣, 等. 广东省鼠形动物携带巴尔通体的调查和基因特征分析[J]. 中国人兽共患病学报, 2018, 34(5): 482-486, 491. Li XY, Chen JD, Li CR, et al. Genotype characteristics of Bartonella and the infection in rodents in Guangdong province, China[J]. Chin J Zoonoses, 2018, 34(5): 482-486, 491. DOI:10.3969/j.issn.1002-2694.2018.00.062 |
| [10] |
姜亚运, 鲁亮, 宋秀平, 等. 宁夏不同栖息环境啮齿动物巴尔通体感染状况调查[J]. 中国媒介生物学及控制杂志, 2016, 27(3): 235-240. Jiang YY, Lu L, Song XP, et al. Investigation of the Bartonella infection in wild rodent populations in Ningxia Hui autonomous region, China[J]. Chin J Vector Biol Control, 2016, 27(3): 235-240. DOI:10.11853/j.issn.1003.8280.2016.03.006 |
| [11] |
宋秀平, 栗冬梅, 贾丽军, 等. 内蒙古小型兽类巴尔通体感染情况调查[J]. 中国媒介生物学及控制杂志, 2015, 26(3): 233-237. Song XP, Li DM, Jia LJ, et al. Investigation of Bartonella infection in small mammals in Inner Mongolia, China[J]. Chin J Vector Biol Control, 2015, 26(3): 233-237. DOI:10.11853/j.issn.1003.4692.2015.03.004 |
| [12] |
Li DM, Hou Y, Song XP, et al. High prevalence and genetic heterogeneity of rodent-borne Bartonella species on Heixiazi Island, China[J]. Appl Environ Microbiol, 2015, 81(23): 7981-7992. DOI:10.1128/AEM.02041-15 |
| [13] |
Paziewska A, Harris PD, Zwolińska L, et al. Recombination within and between species of the Alpha proteobacterium Bartonella infecting rodents[J]. Microb Ecol, 2011, 61(1): 134-145. DOI:10.1007/s00248-010-9735-1 |
| [14] |
Mediannikov O, Ivanov L, Zdanovskaya N, et al. Molecular screening of Bartonella species in rodents from the Russian Far East[J]. Ann N Y Acad Sci, 2005, 1063: 308-311. DOI:10.1196/annals.1355.049 |