 2020, Vol. 31
2020, Vol. 31扩展功能
文章信息
- 相蓉, 郭宪国
- XIANG Rong, GUO Xian-guo
- 二齿新蚤的研究现状
- Current research on Neopsylla bidentatiformis
- 中国媒介生物学及控制杂志, 2020, 31(5): 623-629
- Chin J Vector Biol & Control, 2020, 31(5): 623-629
- 10.11853/j.issn.1003.8280.2020.05.025
-
文章历史
- 收稿日期: 2020-05-23




蚤类是鼠疫和地方性斑疹伤寒等自然疫源性疾病的传播媒介,其最重要的医学意义是传播鼠疫。近年来,鼠疫已在部分地区出现“复燃”和流行回升,2016年马达加斯加发生肺鼠疫并逐年上升,病死率高达22.20%,2019年在蒙古国的巴彦-洛吉亚省有2人因败血症鼠疫导致死亡[1],2019年11月北京朝阳医院确诊2例由内蒙古自治区(内蒙古)输入的肺鼠疫患者[2]。从全球范围来看,鼠疫等蚤媒传染病正在“卷土重来”,其发病率呈上升趋势,这一过程可能与气候变化和环境破坏有关[1]。目前,我国货物及人口流动频繁,我国的鼠疫疫源地面积有不断扩大的趋势,鼠疫流行的潜在风险依然存在,鼠疫防治工作形势仍然很严峻[3]。
蚤类是属于昆虫纲(Insecta)、蚤目(Siphonaptera)的节肢动物。迄今为止,全世界已发现约2 500种(亚种)蚤类,隶属于5总科16科239属,我国已发现有近650种(亚种)蚤类,隶于4总科10科74属[4-5]。其中,新蚤属(Neopsylla)全世界共记录有65种(亚种),我国目前已知分布有46种(亚种),该属有些种类已经被证明可以有效传播鼠疫耶尔森菌(鼠疫菌),有的种类体内存在鼠疫菌的自然感染,医学意义较大。二齿新蚤(N. bidentatiformis)是新蚤属中的一个独立种类,在我国北方地区分布较广并与鼠疫有关,但目前尚未见关于该蚤的详细综述性文献报道。本文在收集他人既往对该蚤系列研究文献的基础上,对二齿新蚤的鉴别特征、与近缘蚤种的区别、地理分布、媒介地位和生态习性等方面的研究现状进行了综述和介绍,以归纳和总结迄今为止对该蚤的研究成果,进而为媒介蚤种及其所传播的蚤媒传染病的预测、预警和防控提供更多的科学依据。
1 二齿新蚤的形态特征及与近缘种关系 1.1 二齿新蚤的鉴别特征雄虫:第9腹板后臂略呈棒形,端部腹缘处只有1列刺形鬃(可分为长短两组)。雌虫:第7背板主鬃列在气门下无鬃以及第7腹板后缘有较长而形如指状后凸之背叶[4]。新蚤属毛新蚤种团中的红羊新蚤(N. hongyangensis)与二齿新蚤是近缘种类,其形态结构的相似度较高,这往往给分类鉴定带来一些困难。传统观点认为,二齿新蚤和红羊新蚤的主要区别在于雄蚤抱器的某些特征:红羊新蚤雄性第9腹板肘部处腹面中央有一向后方伸出的瓶形腹小臂,而二齿新蚤没有[4]。
1.2 二齿新蚤与近缘种的区别由于红羊新蚤与二齿新蚤是近缘蚤种,在分类鉴定中两者成虫(尤其是雌性)通常难以鉴别。李贵真等[6]在红羊新蚤形态特征的研究中,对二齿新蚤和红羊新蚤成虫的形态结构做了比较。见表 1。

|
雄性第9腹板肘部处腹面中央向后方伸出的瓶形腹小臂,是红羊新蚤与二齿新蚤最主要的鉴别特征,根据这一特征,可以区别这2种近缘蚤种。
此外,肖柏林[7]在实验室条件下,对红羊新蚤的幼虫进行了培养,并与二齿新蚤幼虫形态特征进行了比较(表 2),发现两者幼虫之间存在一定的差异。
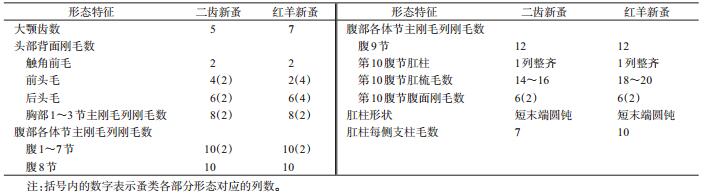
|
国内学者曾经对我国二齿新蚤种团(bidentatiformis group)中7种新蚤的线粒体基因组16S rRNA基因片段差异进行了比较研究,探讨了这7种新蚤的分子系统发育关系[8]。采用CLUSTAL软件排列,认为二齿新蚤和红羊新蚤可以根据变异位点分成2个类群。目前多数研究者认为,二齿新蚤和红羊新蚤是2个不同的蚤种,属于近缘种。但也有研究者认为,二齿新蚤和红羊新蚤可能属于同一种内的2种多态类型。对采自不同地区大量二齿新蚤标本的形态结构进行对比观察后发现:采自北京市和吉林省等地的少部分二齿新蚤标本中,其第9腹板肘部处腹面中央也有1个向后方伸出的瓶形腹小臂状结构,与红羊新蚤类似,但没有红羊新蚤的结构典型,这一发现提示二齿新蚤可能存在种内形态变异[8]。对采自北京市和内蒙古的二齿新蚤和红羊新蚤进行线粒体基因组16S rRNA的序列分析显示,二者之间最大序列差异<1%,认为二齿新蚤和红羊新蚤可能属于同一种内的2种多态类型。进一步的研究发现,瓶形腹小臂状的结构在青海省、宁夏回族自治区(宁夏)和内蒙古等地的二齿新蚤中比较普遍,其发生率远高于北京市和吉林省等华北地区的二齿新蚤[8-9]。根据上述结果,研究者推测瓶形腹小臂状结构可能是由基因型决定的,红羊新蚤可能是二齿新蚤的同种异名,但这种推测还没有得到最后证实,红羊新蚤和二齿新蚤的关系还有待进一步研究,对于腹小臂这一特殊结构的机能和相关形态学问题也值得进一步研究[6]。
近年来,DNA条形码技术已被广泛应用于动物、植物、微生物“隐存种”的发现、残缺标本的鉴定以及宿主动物的识别等。DNA条形码技术是利用有足够变异且容易扩增、相对较短的标准DNA片段,在种内的特异性与种间的多样性基础上建立的一种新的生物身份识别系统,能够对物种进行快速、准确的识别和鉴定,且不受物种发育阶段、性别及个体完整性的影响[10-11]。对二齿新蚤和宽新蚤(N. mana)线粒体DNA中1段900个碱基的基因片段的研究发现,这2种新蚤细胞色素氧化酶亚基Ⅱ基因的CG含量都是33.30%(在昆虫中较高),2种新蚤基因间碱基的变异均以T→C转换为主,且变异率比较低(约为1.80%),这一结果提示二齿新蚤和宽新蚤的亲缘关系很近[12]。
2 地理分布二齿新蚤广泛分布于古北界的东北亚界和中亚亚界,在我国北方的青海、黑龙江、吉林、河北、内蒙古、山西、北京、辽宁、山东、陕西、宁夏、河南等12个省(自治区、直辖市)均有分布[13-28],国外分布于韩国[29]、俄罗斯[30]和以色列等地[31]。该蚤的常见生境有林区、草原、森林草原、荒漠草原、丘陵、高原以及农垦区和城市周围。2011-2012年5月,在青海省共和县开展调查时,经鉴定其中14♂、21 ♀为二齿新蚤[13]。2012年,通过对黑龙江省中俄边境地区3市(县)鼠体蚤类的生态研究,表明二齿新蚤为当地优势种(26.60%)[15]。任清明等[18]通过对东北三省多年来采获的大量标本进行整理鉴定后发现,二齿新蚤在吉林(全省)、黑龙江(全省)和辽宁省(部分地区)均有分布。李俊孚[24]通过3年的连续监测,表明二齿新蚤为白城小型鼠体蚤的优势种。任清明等[16]通过对吉林省延边地区蚤类分布的研究发现二齿新蚤为当地优势种。
3 在鼠疫媒介中的地位在传染病媒介判定中,确定媒介蚤种的依据是多方面的,如:该蚤必须吸人血;在该蚤的自然种群中存在某种病原体(如鼠疫菌)的自然感染,即在其体内能够检测到病原体;在实验室内,该蚤能够人工感染病原体,具有形成菌栓的能力;种群数量较大,一般应是当地的优势蚤种,且种群数量的季节消长与所怀疑传播的疾病流行趋势一致等[14, 28, 32-33]。迄今为止,研究表明二齿新蚤基本符合上述鼠疫媒介的判定标准,是我国鼠疫的媒介蚤之一。
3.1 吸血特性蚤类的吸血行为具有重要的流行病学意义。有效的传播媒介必须具备吸血(特别是吸人血)的能力和较长的寿命。1989年,王身荣和房静[34]用二齿新蚤进行实验性吸血,研究出1种达到国际同类水平的皮膜饲喂装置。早在1982年的报道显示[35],1名男性患者被跳蚤叮咬,两腿出现扁平丘疹和斑丘疹,大者如拇指甲,逐渐变成红色,连成一片,奇痒,经鉴定为二齿新蚤所为。马立名[36]通过实验证明,大部分二齿新蚤在与宿主接触30 min内吸血。1996年马立名[37]用人工饲养的800匹二齿新蚤叮咬了20位志愿者,在温度25 ℃,相对湿度60%~70%的条件下,总吸血率为16.25%(130/800),其中雌性52.30%,雄性47.69%。上述实验证明,二齿新蚤具有吸人血的特性,为二齿新蚤可作为人类疾病的传播媒介提供了生物学证据。
除了能够叮刺和吸食人血外,二齿新蚤的寿命也比较长。马立名[38]发现,新羽化的二齿新蚤在不供给血液条件下也能存活很长或较长时间,羽化后耐饥力较强,如果未成年期和幼虫期的营养供给不足,其生存时间会受到明显影响。该蚤寿命较长的特点对蚤类种群延续和传播疾病具有重要的流行病学意义。
3.2 对鼠疫菌的自然感染、人工感染及其媒介效能在我国多种类型鼠疫自然疫源地〔喜马拉雅旱獭(Marmota himalayana)、达乌尔黄鼠(Spermophilus dauricus)、长爪沙鼠(Meriones unguiculatus)和布氏田鼠(Lasiopodomys brandtii)鼠疫疫源地等〕,都曾从二齿新蚤体内检测到鼠疫菌,即该蚤的自然种群存在鼠疫病原体的自然感染(自然染疫)。在达乌尔黄鼠疫源地,二齿新蚤在黄鼠体表的寄生蚤中占9.65%,在褐家鼠(Rattus norvegicus)体表占2.15%,在褐家鼠巢占48.77%,在小家鼠(Mus musculus)巢占58.33%,在黑线仓鼠(Cricetulus barabensis)和黑线姬鼠(Apodemus agrarius)中分别占90.13 %和83.33% [39]。国内学者通过野外实践调查和实验研究,证实二齿新蚤属高菌栓形成率蚤类(鼠疫实验感染率为86.00%,栓塞率为47.00%),该蚤在感染率、栓塞率、菌栓形成速度和菌栓蚤寿命等方面与印鼠客蚤(Xenopsylla cheopis)(鼠疫的高效传播媒介)均较接近,对鼠疫流行程度的加大起了重要作用[40]。王身荣等[41]在实验室测得二齿新蚤的媒介效能为0.11,媒介指数为0.01。由于二齿新蚤的栓塞率较高,在鼠疫流行期间,该蚤在自然界的实际媒介能力应该会超过实验室测定结果。有学者认为,在我国的多种鼠疫自然疫源地〔喜马拉雅旱獭、阿拉善黄鼠(S. alaschanicus)、长爪沙鼠和布氏田鼠鼠疫疫源地等〕,二齿新蚤可能会把主要宿主动物体内的鼠疫菌播散给其他小型哺乳类(包括家栖鼠类),这对促进不同宿主间的鼠疫传播和加大鼠疫流行强度等,均有重要意义[41]。
3.3 蚤体菌栓的形成判断一个蚤种是否能够成为鼠疫的有效传播媒介,首先要看其前胃是否能够形成菌栓,而菌栓的形成与蚤前胃的形态密切相关[42]。但关于二齿新蚤前胃结构和菌栓形成的报道较少,多数文献是关于红羊新蚤(二齿新蚤的近缘种)前胃结构和菌栓形成的研究报道。红羊新蚤的前胃呈圆灯笼状,前胃刺上齿分布于刺长的下1/3处,齿小而稀,蚤的前胃刺可以防止中肠中已存的血液向食道和咽部逆流。红羊新蚤前胃刺基部立体感较强呈矛形,尾端形态有梭镖形、吸管形;每刺均有较深的凹槽,且形态多样;前胃刺的凹槽和齿,适宜血液中有形成分的黏附和沉积,利于细菌在此处堆积和孳生,使菌栓形成成为可能[43]。通过二齿新蚤和红羊新蚤的对比发现,二者在前胃形态构造上无差异,所以二齿新蚤的前胃也应该可以形成菌栓[5-7],进一步证实了二齿新蚤可以形成菌栓,能够成为传播鼠疫的有效传播媒介[41, 43-44]。
4 生态习性 4.1 季节分布二齿新蚤的季节消长因地理区域的不同而存在一定变化,这可能与多种因素有关,如媒介蚤种本身的生活习性、宿主的活动规律以及不同地区环境温度的差异等[27-28]。我国陕西省的调查结果显示,2004年长爪沙鼠体表寄生的二齿新蚤季节消长只有8月1个高峰,而2007年黑线仓鼠体表寄生的二齿新蚤在4和9月出现双峰曲线[28]。刘国平等[45]通过对我国东北中朝俄边境地区鼠体外寄生虫重要种群的季节消长调查,发现二齿新蚤在4月下旬和6月上旬出现2次高峰。李维贤等[46]通过对辽宁省西北部的阜新、建平县进行调查,发现二齿新蚤作为当地优势种,在5和9月出现2个高峰期,这些与陕西省在2004和2007年的观测情况不同。Bitam等[47]认为,对于很多蚤类,温度和湿度是影响其生存和发展至关重要的因素。由于气候变化的频繁和生态环境的破坏,影响了蚤类的发育、分布及相关参数。来自吉林省鼠疫自然疫源地的监测结果显示,原有鼠疫自然疫源地的蚤种由43种减少为14种,二齿新蚤的数量相对增加(达总蚤数的7.54%),这可能是近几年人类活动对原有生态环境的干扰或破坏(如耕地不断扩展、兴修水利、植树造林等)的结果[48]。由于人为干扰和破坏,原有宿主动物的生存环境受到了不同程度的影响,栖息地面积的减少可能导致宿主种类和数量的异常变化。通常,部分寡宿主型的蚤种对宿主的选择和寄生比较严格,宿主种类和数量的异常变化可能使其无法生存,导致其数量减少或消失,这样就间接导致了二齿新蚤数量比例的增加。因此,研究不同鼠疫自然疫源地媒介蚤种的种类构成和季节消长,对于揭示鼠疫流行规律和有效预防鼠疫均具有重要意义。
4.2 取食习性与环境因素关系与其他绝大多数的蚤种类似,二齿新蚤雌性和雄性成虫都吸血。蚤的幼虫主要生活在宿主动物的窝巢,口器是咀嚼式口器,杂食性,不吸血,但经常取食成虫排出的血便[5]。马立名[49]用吸血后二齿新蚤的中肠血液占据腹节数的方法进行观察,发现羽化24 h以上的“老龄成蚤”(原文直接称为“成蚤”)的吸血量大于羽化24 h以内的“青年成蚤”(原文错误地称为“幼蚤”)(P<0.05,方差分析),雌蚤吸血量大于雄蚤(P<0.01,方差分析)。青年成蚤在温度为25 ℃时吸血率最高;老龄成蚤在各种温度下的吸血率均接近100 %。蚤吸血量与吸血时的温度无关,高温时蚤的血液消化速度较低温时快。在温度25 ℃以下时,老龄成蚤消化速度较青年成蚤快;30 ℃以上时,二者消化速度相近,消化速度与性别无关。
二齿新蚤既侵袭活寄主,也侵袭死寄主,侵袭蚤数均很多。活寄主体蚤数与环境温度呈直线负相关,死寄主体蚤数与环境温度不相关。在环境温度25 ℃以下时,活寄主体蚤数多于死寄主体,35 ℃以上时,活寄主体蚤数少于死寄主体[50]。1985-1990年,马立名[51-53]进行了一系列实验,通过设置温度、湿度、吸血次数和性别条件,观察二齿新蚤在不同条件下的存活力。通过统计学方法,不仅描述了二齿新蚤在不同水平下的平均存活日数,还得出了关于二齿新蚤性别的方差分析结果。李仲来和马立明[54-55]曾利用多种统计学方法(折线回归、4种双因素方差分析和多元线性回归模型)对二齿新蚤的存活力进行了研究。马巧云和李天群[56]在此基础上又进行了进一步的研究和探索,通过生存分析、建立对数线性回归模型和危险率模型等,得到了以下结果:二齿新蚤的生存危险率与温度的变化成正比,与湿度的变化成反比,高温、低湿对蚤类(包括二齿新蚤)的生存不利,低温、高湿对蚤类的生存有利,且温度的影响力大于湿度。另外,吸血条件的改善也会降低蚤类生存到某天的危险率,这一结果与Bitam等[47]的观点相匹配,即:温度和湿度是影响蚤类生存发展至关重要的因素。
王身荣和房静[57]在25 ℃,相对湿度为85%的实验室条件下,用称重法测得刚羽化的二齿新蚤平均吸血量为0.03 μl,其中,雄性0.02 μl,雌性0.03 μl。5日龄后,吸血量达到稳定量0.08 μl,雄性0.07 μl,雌性0.08 μl,此结果与马立名[49]发现的成蚤吸血特点相一致。
4.3 宿主体表及窝巢的分布特点马立名[58]以小白鼠做实验宿主,将650匹二齿新蚤放置在小白鼠腹部中间,1 h后发现二齿新蚤在前腹部占2.62%,前背部占6.26%,后腹部占15.08%,后背部占75.69%,差异有统计学意义(P<0.01)。采用相同的实验方法,以大仓鼠(Tscherskia triton)寄生做野外调查发现,二齿新蚤在前腹部分布最少,前背部稍多,后腹部较多,后背部最多。实验认为,宿主毛层对跳蚤在宿主体表的分布起主要作用,宿主本身的清蚤能力起次要作用。
费荣中等[59]通过对达乌尔黄鼠洞内蚤的空间分布研究发现,二齿新蚤在空间分布上,巢内多于洞干,洞干多于洞口。黎占海等[40]通过对盐池县动物鼠疫流行病的调查发现,在检测到的3 771匹长爪沙鼠体蚤中,二齿新蚤占6.82%;在检测到的329匹长爪沙鼠巢蚤中,二齿新蚤占34.90%,根据这一特征,可将二齿新蚤归属为“巢型蚤”。
5 二齿新蚤雄性阉割畸形要点国内学者将雄蚤外生殖器发育不全或退化的称为“雄性阉割畸形”[60]。王身荣和刘晓倩[60]、黄贵萍和谢音凡[61]通过对新蚤属形态的鉴定,在所保留的蚤类标本中,发现有6种新蚤属、25例亚种存在雄性阉割畸形,其中二齿新蚤9例。在9例二齿新蚤雄性阉割畸形中,有4例内阳茎缩短,阳茎内突骨化较弱,稍缩小。柄突轻度变形,仅末半段超出第8背板前缘,而正常标本末2/3段超出。有5例内阳茎和阳茎内突严重退化且变形,阳茎端结构混乱仅阳茎钩突尚可辨认。第9腹节诸结构缩小,其中柄突除明显缩短外变形亦严重,其末端不达第8腹节前缘或仅稍超过之,第9腹板后臂端部腹缘的2组刺形鬃有的成为1组。第8背板前缘平滑无齿状前凸,第8腹板前缘的指状突退化[60]。由此可以推测,二齿新蚤发生畸形的可能性较大。
综上所述,二齿新蚤分布较为广泛,且宿主较多,在多个自然鼠疫疫源地发现其自然感染鼠疫菌,是鼠疫的传播媒介之一。根据目前的文献分析,发现国外对于二齿新蚤的研究相对较少,国内的相关文献大多年代久远。目前,认为该蚤的研究还存在以下主要问题:(1)二齿新蚤是否存在种内形态变异,其与近缘种红羊新蚤之间的分类地位和关系存在的争议有待进一步澄清。(2)二齿新蚤与其他近缘蚤的系统发育关系及其分子遗传学方面还有待进一步研究和探讨。
| [1] |
龚震宇. 2019年全球鼠疫疫情概况[J]. 疾病监测, 2019, 34(8): 775-776. Gong ZY. General situation of plague in 2019[J]. Dis Surveill, 2019, 34(8): 775-776. DOI:10.3784/j.issn.1003-9961.2019.08.021 |
| [2] |
张怡, 张昊澄. 鼠疫的发病机制研究进展[J]. 微生物与感染, 2019, 14(6): 326-332. Zhang Y, Zhang HC. Advances in research on the pathogenesis of plague[J]. J Microbes Infect, 2019, 14(6): 326-332. DOI:10.3969/j.issn.1673-6184.2019.06.002 |
| [3] |
梓豪. 鼠疫的传播途径有哪些?如何预防?[J]. 智慧中国, 2019(11): 76-78. Zi H. What are the transmission ways of plague? How to prevent it?[J]. Wisdom China, 2019(11): 76-78. |
| [4] |
中国科学院中国动物志编辑委员会. 中国动物志[M]. 北京: 科学出版社, 2007: 47-527. Editorial Committee of Zoology of China, Chinese Academy of Sciences. Fauna sinica[M]. Beijing: Science Press, 2007: 47-527. |
| [5] |
解宝琦, 曾静凡. 云南蚤类志[M]. 昆明: 云南科技出版社, 2001: 1-28. Xie BQ, Zeng JF. The siphonaptera of Yunnan[M]. Kunming: Yunnan Science and Technology Press, 2001: 1-28. |
| [6] |
李贵真, 吴文贞, 张荣广. 红羊新蚤的地理分布、宿主关系和鉴别特征的研究(蚤目:多毛蚤科)[J]. 昆虫学报, 1995, 38(2): 161-165. Li GZ, Wu WZ, Zhang RG. On the geographical distribution, host preference and the diagnostic characters of Neopsylla hongyangensis Li, Bai & Chen (Siphonaptera:hystrichopsyllidae)[J]. Acta Entomol Sin, 1995, 38(2): 161-165. DOI:10.16380/j.kcxb.1995.02.006 |
| [7] |
肖柏林. 红羊新蚤的幼虫形态及与二齿新蚤幼虫的比较[J]. 昆虫学报, 1993, 36(1): 67-70. Xiao BL. Morphological observation of the larvae of Neopsylla hongyangensis and their comparison with larvae of N. bidentatiformis[J]. Acta Entomol Sin, 1993, 36(1): 67-70. DOI:10.16380/j.kcxb.1993.01.012 |
| [8] |
鲁亮, 吴厚永. 基于16S rRNA序列的新蚤属二齿新蚤种团部分种类的分子系统发育关系[J]. 昆虫学报, 2001, 44(4): 548-554. Lu L, Wu HY. The molecular phylogeny of some species of the bidentatiformis group of the genus Neopsylla based on 16S rRNA gene[J]. Acta Entomol Sin, 2001, 44(4): 548-554. DOI:10.16380/j.kcxb.2001.04.024 |
| [9] |
李国军, 李帅一, 石杲. 内蒙古新蚤属的种类及鼠疫流行病学意义探讨[J]. 疾病监测与控制, 2014, 8(4): 241-242. Li GJ, Li SY, Shi G. An analysis of types of Neopsylla genus and the meaning of the plague epidemiology in Inner Mongolia[J]. J Dis Monit Control, 2014, 8(4): 241-242. |
| [10] |
张瑞玲, 王晓旭, 张忠. DNA条形码技术在医学媒介生物鉴定中的应用[J]. 中国热带医学, 2017, 17(8): 831-837. Zhang RL, Wang XX, Zhang Z. The application of DNA barcode in the identification of medical vectors[J]. China Trop Med, 2017, 17(8): 831-837. DOI:10.13604/j.cnki.46-1064/r.2017.08.24 |
| [11] |
杨倩倩, 刘苏汶, 俞晓平. DNA条形码分析方法研究进展[J]. 应用生态学报, 2018, 29(3): 1006-1014. Yang QQ, Liu SW, Yu XP. Research progress on DNA barcoding analysis methods[J]. Chin J Appl Ecol, 2018, 29(3): 1006-1014. DOI:10.13287/j.1001-9332.201803.032 |
| [12] |
鲁亮, 吴厚永. 两种新蚤细胞色素氧化酶亚基Ⅱ基因的克隆和测序[J]. 寄生虫与医学昆虫学报, 2000, 7(3): 170-174. Lu L, Wu HY. Cloning and sequence analyses of COⅡ gene of two Neopsylla species[J]. Acta Parasitol Med Entomol Sin, 2000, 7(3): 170-174. DOI:10.3969/j.issn.1005-0507.2000.03.007 |
| [13] |
郑谊, 马英, 罗军, 等. 青海省蚤类一新纪录[J]. 中国媒介生物学及控制杂志, 2013, 24(3): 239-240. Zheng Y, Ma Y, Luo J, et al. A new record of flea species in Qinghai province, China[J]. Chin J Vector Biol Control, 2013, 24(3): 239-240. |
| [14] |
吴海生, 吴海莲, 杨晓艳, 等. 青海省海北藏族自治州鼠疫耶尔森菌病原学分析及流行病学意义[J]. 中国媒介生物学及控制杂志, 2019, 30(1): 35-39. Wu HS, Wu HL, Yang XY, et al. An etiological study and epidemiological significance of Yersinia pestis in Haibei Tibetan autonomous prefecture, Qinghai province, China[J]. Chin J Vector Biol Control, 2019, 30(1): 35-39. DOI:10.11853/j.issn.1003.8280.2019.01.007 |
| [15] |
任清明, 王峰, 王旭, 等. 黑龙江省中俄边境地区鼠体蚤类的生态研究[J]. 中华卫生杀虫药械, 2013, 19(1): 56-58. Qen QM, Wang F, Wang X, et al. Ecological study of fleas on the ecotparasites of the rodent in frontiers of Russia and Heilongjiang province[J]. Chin J Hyg Insect Equip, 2013, 19(1): 56-58. DOI:10.19821/j.1671-2781.2013.01.017 |
| [16] |
任清明, 王峰, 刘国平, 等. 吉林省延边地区蚤类分布与季节消长的研究[J]. 医学动物防制, 2020, 36(3): 205-207. Ren QM, Wang F, Liu GP, et al. Studies on the distribution and seasonal fluctuation of fleas in Yanbian area of Jilin province[J]. J Med Pest Control, 2020, 36(3): 205-207. DOI:10.7629/yxdwfz202003001 |
| [17] |
姚晓恒, 李长华, 刘铁于, 等. 2008-2018年吉林省乾安县鼠疫监测数据分析[J]. 中国地方病防治杂志, 2019, 34(4): 417-418. Yao XH, Li CH, Liu TY, et al. Analysis of plague monitoring data in Qian'an country, Jilin province 2008-2018[J]. Chin J Control Endem Dis, 2019, 34(4): 417-418. |
| [18] |
任清明, 王旭, 刘国平, 等. 我国东北三省蚤的种类和区系分布研究[J]. 中华卫生杀虫药械, 2019, 25(3): 266-272. Ren QM, Wang X, Liu GP, et al. Species and fauna distribution of fleas in three provinces of northeast China[J]. Chin J Hyg Insect Equip, 2019, 25(3): 266-272. DOI:10.19821/j.1671-2781.2019.03.021 |
| [19] |
武大胜, 刘俊平, 吕少军. 康保县2013-2015年蚤类监测数据分析[J]. 中国地方病防治杂志, 2016, 31(3): 287. Wu DS, Liu JP, Lyu SJ. Analysis of flea monitoring data in Kangbao country from 2013 to 2015[J]. Chin J Control Endem Dis, 2016, 31(3): 287. |
| [20] |
魏怀波, 杨喜凤, 田丽, 等. 2006-2016年内蒙古边境口岸蚤类分布[J]. 中国国境卫生检疫杂志, 2019, 42(3): 175-178, 192. Wei HB, Yang XF, Tian L, et al. Distribution of fleas in the frontier ports of Inner Mongolia, 2006-2016[J]. Chin J Front Health Quarantine, 2019, 42(3): 175-178, 192. DOI:10.16408/j.1004-9770.2019.03.007 |
| [21] |
吴美和, 赵亮怀. 山西省天镇县1987-2010年鼠疫监测结果[J]. 中国地方病防治杂志, 2011, 26(6): 448-449. Wu MH, Zhao LH. Plague surveillance results in Tianzhen county, Shanxi province, 1987-2010[J]. Chin J Control Endem Dis, 2011, 26(6): 448-449. |
| [22] |
郭天宇, 许荣满. 北京东灵山地区鼠类体外寄生虫时间生态位的研究[J]. 中国媒介生物学及控制杂志, 2003, 14(1): 30-32. Guo TY, Xu RM. Study on time niche among the ectoparasites of the rodent in Dongling Mountain in Beijing[J]. Chin J Vector Biol Control, 2003, 14(1): 30-32. |
| [23] |
周光智, 程绪浩, 韩珂嘉, 等. 某部营区蚤侵害调查及灭蚤效果观察[J]. 中华卫生杀虫药械, 2016, 22(6): 556-558. Zhou GZ, Cheng XH, Han KJ, et al. Infestation investigation and evaluation of fleas control effects in a military camp[J]. Chin J Hyg Insect Equip, 2016, 22(6): 556-558. DOI:10.19821/j.1671-2781.2016.06.011 |
| [24] |
李俊孚. 2014-2016年白城市鼠疫监测结果分析[J]. 中国卫生产业, 2017, 14(8): 25-26. Li JF. Analysis of plague monitoring results in Baicheng city from 2014 to 2016[J]. China Health Ind, 2017, 14(8): 25-26. DOI:10.16659/j.cnki.1672-5654.2017.08.025 |
| [25] |
白学礼. 宁夏二齿新蚤和红羊新蚤的地理分布及其与宿主关系的研究[J]. 疾病预防控制通报, 2015, 30(5): 38-40. Bai XL. Study on geographical distribution of Neopsylla bidentatiformis and N. hongyangensis in Ningxia and relationship with their hosts[J]. Bull Dis Control Prev, 2015, 30(5): 38-40. DOI:10.13215/j.cnki.jbyfkztb.1411032 |
| [26] |
白学礼. 宁夏蚤类区系及其分布特征[J]. 寄生虫与医学昆虫学报, 2019, 26(1): 47-58. Bai XL. Flea fauna and distribution in Ningxia[J]. Acta Parasitol Med Entomol Sin, 2019, 26(1): 47-58. |
| [27] |
孙养信, 吕永杰, 白江春, 等. 陕西省鼠疫疫区媒介蚤季节消长研究[J]. 中国媒介生物学及控制杂志, 2005, 16(6): 442-445. Sun YX, Lyu YJ, Bai JC, et al. Study on the seasonal variation of vector fleas in plague epidemic area, Shaanxi province[J]. Chin J Vector Biol Control, 2005, 16(6): 442-445. |
| [28] |
孙养信, 阮春来, 白江春, 等. 陕西省鼠疫疫区夜行鼠寄生蚤调查[J]. 中国病原生物学杂志, 2008, 3(2): 133-135. Sun YX, Ruan CL, Bai JC, et al. Investigation on the parasitic fleas of nocturnal rodents in plague epidemic area, Shaanxi province[J]. J Pathog Biol, 2008, 3(2): 133-135. DOI:10.3969/j.issn.1673-5234.2008.02.015 |
| [29] |
Kim BJ, Kim SJ, Kang JG, et al. First report for the seasonal and annual prevalence of flea-borne Bartonella from rodents and soricomorphs in the Republic of Korea[J]. Vector Borne Zoonotic Dis, 2013, 13(7): 457-467. DOI:10.1089/vbz.2012.1115 |
| [30] |
Medvedev SG, Dokuchaev NT, Tretyakov KA, et al. Fleas of small mammals from the North of the Russian Far East[J]. Entomol Rev, 2014, 94(9): 1288-1296. DOI:10.1134/S0013873814090097 |
| [31] |
Krasnov BR, Shenbrot GI, Khokhlova IS, et al. Sex ratio in flea infrapopulations:number of fleas, host gender and host age do not have an effect[J]. Parasitology, 2008, 135(9): 1133-1141. DOI:10.1017/S0031182008004551 |
| [32] |
寻慧, 漆一鸣. 媒介鼠蚤与鼠疫关系的研究进展[J]. 贵阳医学院学报, 2001, 26(4): 318-320. Xun H, Qi YM. Research progress on the relationship between rodent fleas and plague[J]. J Guiyang Med Coll, 2001, 26(4): 318-320. DOI:10.19367/j.cnki.1000-2707.2001.04.015 |
| [33] |
王祖郧, 魏柏青, 张珊瑚, 等. 中国自然感染鼠疫杆菌的蚤、蜱、螨与虱类[J]. 中国人兽共患病学报, 2007, 23(3): 307-309. Wang ZY, Wei BQ, Zhang SH, et al. Fleas, ticks, mites and ticks naturally infected with Yersinia pestis in China[J]. Chin J Zoonoses, 2007, 23(3): 307-309. DOI:10.3969/j.issn.1002-2694.2007.03.030 |
| [34] |
王身荣, 房静. 用于蚤类吸血研究的一种皮膜饲喂装置[J]. 昆虫知识, 1989, 26(3): 175-177. Wang SR, Fang J. A kind of skin film feeding device for the study of flea blood absorption[J]. Chin J Appl Entomol, 1989, 26(3): 175-177. |
| [35] |
马立名, 戴钦英, 陈晶杰, 等. 野鼠寄生蚤叮人两例[J]. 中国寄生虫学与寄生虫病杂志, 1991, 9(1): 70. Ma LM, Dai QY, Chen JJ, et al. Two cases of human bited by parasitic flea of wild rat[J]. Chin J Parasitol Parasit Dis, 1991, 9(1): 70. |
| [36] |
马立名. 二齿新蚤和方形黄鼠蚤松江亚种吸血习性的实验室观察[J]. 四川动物, 1992, 11(1): 13-15. Ma LM. Laboratory observation on the blood sucking habits of Neopsylla bidentatiformis and Citellophilus tesquorum sungaris[J]. Sichuan J Zool, 1992, 11(1): 13-15. |
| [37] |
马立名. 二齿新蚤叮吸人血的几项试验[J]. 昆虫知识, 1996, 33(2): 102-103. Ma LM. Several experiments on human blood sucking by Neopsylla bidentatiformis[J]. Entomol Knowl, 1996, 33(2): 102-103. |
| [38] |
马立名. 两种未吸血幼蚤的耐饥力[J]. 四川动物, 1991, 10(4): 12-14. Ma LM. Starvation tolerance of two kinds of fleas without sucking blood[J]. Sichuan J Zool, 1991, 10(4): 12-14. |
| [39] |
王身荣, 樊振亚, 马立名, 等. 达乌尔黄鼠疫源地几种蚤的鼠疫流行病学意义[J]. 中国地方病防治杂志, 1994, 9(2): 111-112. Wang SR, Fan ZY, Ma LM, et al. Epidemiological significance of several fleas in Citellus dauricus[J]. Chin J Control Endem Dis, 1994, 9(2): 111-112. |
| [40] |
黎占海, 李生起, 郭臻, 等. 盐池县1987年动物鼠疫流行病学调查报告[J]. 中国地方病防治杂志, 1989, 4(2): 117-118. Li ZH, Li SQ, Guo Z, et al. An epidemiological survey of animal plague in Yanchi county in 1987[J]. Chin J Control Endem Dis, 1989, 4(2): 117-118. |
| [41] |
王身荣, 徐诚, 刘晓倩, 等. 二齿新蚤与印鼠客蚤鼠疫媒介效能的比较研究[J]. 中国地方病学杂志, 1993, 12(6): 348-350. Wang SR, Xu C, Liu XQ, et al. Studies on comparative plague-vector efficiency of Neopsylla bidentatiformis and Xenopsylla cheopis[J]. Chin J Endemiol, 1993, 12(6): 348-350. |
| [42] |
刘哲, 郭宪国, 杨志华. 斧形盖蚤的研究现状[J]. 中国媒介生物学及控制杂志, 2018, 29(6): 662-665. Liu Z, Guo XG, Yang ZH. Overview of research on Callopsylla dolabris[J]. Chin J Vector Biol Control, 2018, 29(6): 662-665. DOI:10.11853/j.issn.1003.8280.2018.06.030 |
| [43] |
张全芬, 杨锡正, 马英, 等. 青海不同鼠疫自然疫源地内7种蚤前胃形态结构光镜研究[J]. 地方病通报, 2003, 18(4): 15-19. Zhang QF, Yang XZ, Ma Y, et al. Microscopic study on shape and structure of the proventriculi of seven species of fleas from plague natural foci in Qinghai[J]. Endem Dis Bull, 2003, 18(4): 15-19. DOI:10.3969/j.issn.1000-3711.2003.04.005 |
| [44] |
马英, 杨锡正, 张全芬. 青海省不同鼠疫疫源地内7种媒介蚤前胃形态结构Ⅲ.扫描电镜研究[J]. 中国人兽共患病杂志, 2004, 20(3): 266-267. Ma Y, Yang XZ, Zhang QF. Morphological structure of the proventriculi of seven species of fleas from plague natural foci in Qinghai province Ⅲ. Scanning electron microscope study[J]. Chin J Zoonoses, 2004, 20(3): 266-267. DOI:10.3969/j.issn.1002-2694.2004.03.033 |
| [45] |
刘国平, 周旭, 任清明, 等. 边境地区鼠体外寄生虫重要种群的季节消长[J]. 中华卫生杀虫药械, 2007, 13(6): 444-446. Liu GP, Zhou X, Ren QM, et al. Seasonal fluctuation of predominant species on the ectoparasites of the rodent[J]. Chin J Hyg Insect Equip, 2007, 13(6): 444-446. DOI:10.3969/j.issn.1671-2781.2007.06.018 |
| [46] |
李维贤, 张放, 潘凤纯, 等. 辽宁西部地区褐家鼠寄生蚤调查研究[J]. 中国媒介生物学及控制杂志, 1993, 4(4): 279-281. Li WX, Zhang F, Pan FC, et al. Studies on the parasitic fleas at Rattus norvegicus in the west of Liaoning province[J]. Chin J Vector Biol Control, 1993, 4(4): 279-281. |
| [47] |
Bitam I, Dittmar K, Parola P, et al. Fleas and flea-borne diseases[J]. Int J Infect Dis, 2010, 14(8): e667-e6676. DOI:10.1016/j.ijid.2009.11.011 |
| [48] |
张贵, 张芳, 刘振才, 等. 吉林省鼠疫自然疫源地蚤类群落结构的研究[J]. 中国媒介生物学及控制杂志, 2007, 18(2): 124-126. Zhang G, Zhang F, Liu ZC, et al. Study of fleas community structures on Jilin plague natural focus[J]. Chin J Vector Biol Control, 2007, 18(2): 124-126. DOI:10.3969/j.issn.1003-4692.2007.02.014 |
| [49] |
马立名. 二齿新蚤的吸血活动[J]. 昆虫学报, 1985, 28(2): 188-194. Ma LM. The blood-sucking activity of Neopsylla bidentatiformis[J]. Acta Entomol Sin, 1985, 28(2): 188-194. DOI:10.16380/j.kcxb.1985.02.011 |
| [50] |
马立名. 北方几种蚤类侵袭寄主的研究和斧形盖蚤叮人试验[J]. 生态学报, 1987, 7(2): 154-160. Ma LM. Studies on the attacking behaviors of some species of flea and experiment on Callopsylla dolabris bitting man[J]. Acta Ecol Sin, 1987, 7(2): 154-160. |
| [51] |
马立名. 二齿新蚤和方形黄鼠蚤松江亚种成虫在不同条件下寿命的观察[J]. 昆虫知识, 1990, 27(6): 358-359. Ma LM. Observation the life span under different conditions of the adults of fleas Citellophilus tesquorum sungaris and Neopsylla bidentatiformis[J]. Chin J Appl Entomol, 1990, 27(6): 358-359. |
| [52] |
马立名. 二齿新蚤和方形黄鼠蚤松江亚种的耐饥力[J]. 中国媒介生物学及控制杂志, 1993, 4(5): 350-354. Ma LM. Hunger resistance of fleas Neopsylla bidentatiformis and Citellophilus tesquorum sungaris[J]. Chin J Vector Biol Control, 1993, 4(5): 350-354. |
| [53] |
马立名. 二齿新蚤和方形黄鼠蚤松江亚种成虫寿命与温湿度的关系[J]. 地方病通报, 1992, 7(4): 89-91. Ma LM. Life duration of adult Neopsylla bidentatiformis and Citellophtilus tesquorum sungaris in relation to temperature and humidity[J]. Endem Dis Bull, 1992, 7(4): 89-91. DOI:10.13215/j.cnki.jbyfkztb.1992.04.029 |
| [54] |
李仲来, 马立名. 二齿新蚤和方形黄鼠蚤松江亚种存活力的进一步研究[J]. 寄生虫与医学昆虫学报, 1996, 3(1): 44-49. Li ZL, Ma LM. Further studies on survival of fleas Neopsylla bidentatiformis and Citellophilus tesquorum sungaris[J]. Acta Parasitol Med Entomol Sin, 1996, 3(1): 44-49. |
| [55] |
李仲来, 马立名. 二齿新蚤和方形黄鼠蚤松江亚种存活力的再次研究[J]. 寄生虫与医学昆虫学报, 1998, 5(3): 174-178. Li ZL, Ma LM. Restudy on survivorship of the fleas Neopsylla bidentatiformis and Citellophilus tesquorum sungaris[J]. Acta Parasitol Med Entomol Sin, 1998, 5(3): 174-178. |
| [56] |
马巧云, 李天群. 二齿新蚤的对数线性回归模型[J]. 湖北民族学院学报:自然科学版, 2001, 19(2): 51-53. Ma QY, Li TQ. The log-linear regression models of flea Neopsylla bidentiformis[J]. J Hubei Inst Nationalities:Nat Sci, 2001, 19(2): 51-53. DOI:10.3969/j.issn.1008-8423.2001.02.016 |
| [57] |
王身荣, 房静. 二齿新蚤吸血量的测定[J]. 中国地方病防治杂志, 1990, 5(4): 212-213. Wang SR, Fang J. Determination of blood absorption of flea Neopsylla bidentatiformis[J]. Chin J Control Endem Dis, 1990, 5(4): 212-213. |
| [58] |
马立名. 我国北方蚤类在宿主体表的分布[J]. 昆虫学报, 1983, 26(4): 409-412. Ma LM. Distribution of fleas in the hair coat of the host[J]. Acta Entomol Sin, 1983, 26(4): 409-412. DOI:10.16380/j.kcxb.1983.04.008 |
| [59] |
费荣中, 李景原, 徐宝娟, 等. 达乌尔黄鼠洞内蚤的空间分布[J]. 昆虫学报, 1981, 24(4): 397-402. Fei RZ, Li JY, Xu BJ, et al. On the spatial distribution of the fleas in burrows of Citellus dauricus in inner mongol, China[J]. Acta Entomol Sin, 1981, 24(4): 397-402. DOI:10.16380/j.kcxb.1981.04.009 |
| [60] |
王身荣, 刘晓倩. 我国六种新蚤雄性阉割描记及关于单齿新蚤的商榷(蚤目:多毛蚤科)[J]. 昆虫学报, 1993, 36(2): 225-230. Wang SR, Liu XQ. Castrated males in genus Neopsylla from China and a discussion about N. monodentatiformis(Siphonaptera:hystrichopsyllidae)[J]. Acta Entomol Sin, 1993, 36(2): 225-230. DOI:10.16380/j.kcxb.1993.02.019 |
| [61] |
黄贵萍, 谢音凡. 我国东北地区新蚤属一新种记述及有关问题讨论[J]. 四川动物, 1988(1): 9-12. Huang GP, Xie YF. A new species of Neopsylla Wangner, form northeast China, with discussion[J]. Sichuan J Zool, 1988(1): 9-12. |