 2019, Vol. 30
2019, Vol. 30扩展功能
文章信息
- 蔡文凤, 罗剑龙, 苏丽琼, 张四祥, 李寿芹
- CAI Wen-feng, LUO Jian-long, SU Li-qiong, ZHANG Si-xiang, LI Shou-qin
- 云南省剑川县鼠疫疫源地小型兽类体蚤构成及群落多样性特征
- Composition and community diversity of small mammals fleas in the plague foci in Jianchuan county, Yunnan province, China
- 中国媒介生物学及控制杂志, 2019, 30(5): 557-560
- Chin J Vector Biol & Control, 2019, 30(5): 557-560
- 10.11853/j.issn.1003.8280.2019.05.018
-
文章历史
- 收稿日期: 2019-06-18
- 网络出版时间: 2019-08-07 07:00



2 大理白族自治州疾病预防控制中心, 云南 大理 671000;
3 剑川县地方病防治站, 云南 剑川 611300
2 Dali Prefecture Center for Disease Control and Prevention;
3 Jianchuan Center for Endemic Diseases Control
云南省剑川县野鼠鼠疫疫源地位于99°28′~99°56′E,26°10′~ 26°45′N之间,属横断山脉中部,地貌为高山纵谷。鼠疫是人类感染鼠疫耶尔森菌(鼠疫菌)后引起的发病急、死亡率高、传播快的烈性传染病。啮齿动物是鼠疫的主要宿主,其体表寄生蚤(鼠体蚤)是主要传播媒介。1894年,经我国香港地区传播到世界32个国家的第3次世界鼠疫大流行,导致1 500万人死亡[1]。剑川县自1898-1954年间有4年次人间鼠疫流行,1954年发生人间鼠疫34例,死亡15例。1975年7月从沙溪乡石龙村1只自毙的中华姬鼠(Apodemus drara)中分离到鼠疫菌,证实剑川县野鼠鼠疫疫源地的存在[2]。1975-1990年鼠疫流行较为严重,2017年5月,剑川县间隔19年后,再次发生鼠间鼠疫流行,1975年以来,该疫源地鼠疫疫情未波及人间。
蚤类是鼠疫的主要传播媒介,剑川县鼠疫疫源地在1976-2017年鼠疫监测中,从鼠体蚤共分离到鼠疫菌43株[2-3]。小型兽类的体表寄生蚤,随宿主不同会有较大差别,随着主要传播媒介跳蚤种类的不同,防治措施也随之不同。为进一步明确剑川县鼠疫疫源地主要媒介蚤的特征,为制定该鼠疫疫源地的防控措施提供科学依据,我们对2008-2017年的鼠疫监测资料进行了统计分析,现将小型兽类鼠体蚤的构成及群落多样性特征结果报告如下。
1 材料与方法 1.1 资料来源收集剑川县鼠疫疫源地2008-2017年小型兽类体蚤监测资料。剑川县是国家级鼠疫监测点,根据《全国鼠疫监测方案及实施细则》和云南省、大理白族自治州和剑川县的要求,选择剑川县石龙(自然村)为鼠疫固定监测点,每月监测1次,内容包括小型兽类及其体表寄生蚤的监测、室内地面游离蚤和耕地鼠巢蚤监测;流动监测点设17个(自然村),每月选取2个监测点,内容包括小型兽类及其体表寄生蚤的监测;踏查点设46个(自然村),主要是对自毙鼠的搜寻。室内采用笼夜法,园地、耕地、灌木丛采用夹夜法捕获小型兽类,每个生境布放200~250个笼(夹)。
1.2 方法将捕获的小型兽类装入布制鼠袋,带回实验室后,用乙醚麻醉,梳捡体表寄生蚤,将体蚤放在1/10万的龙胆紫生理盐水中保存,1只小型兽类的体蚤1瓶,并编号登记,根据形态学特征,在显微镜下对体蚤进行分类鉴定。
1.3 染蚤率及蚤指数的计算
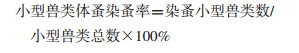
|

|
对体蚤的群落多样性指标采用以下方法测定。
1.4.1 丰富度指数采用Margalef[4]介绍的方法,计算公式:R=(S-1)/lnN,式中,R为群落物种丰富度指数,S为物种数,N为群落中所有物种个体数。R值是指一个群落或某一空间单位的物种数量。
1.4.2 群落多样性指数[5]采用Shannon-Wiener指数,H=-ΣPilnPi(Pi=ni/N),式中,H为多样性指数,Pi为第i种个体在群落中的比例。H值是指一个群落或某一空间单位的物种数量,以及各个物种个体数量的均匀度。
1.4.3 均匀性指数[5]采用Pielou指数,J=H/Hmax,Hmax=lnS,式中,J为均匀性指数,H为多样性指数,S为物种数。J值是指群落组成各物种数量分布的均匀程度。
1.4.4 生态优势度指数[5]采用Simpson指数,D=Σ(Pi)2,式中,D为生态优势度指数,Pi为第i种个体在群落中的比例。D值是反映一个群落中各物种种群数量变化的指标,D值越大,说明群落中物种数量分布越不均匀。
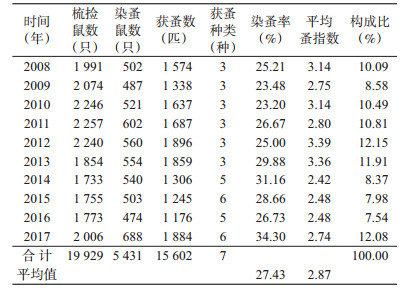
2 结果 2.1 体蚤的染蚤率、平均体蚤指数和年度蚤构成比2008-2017年共梳捡小型兽类19 929只,获体蚤15 602匹,染蚤率在23.20%~34.30%之间,平均染蚤率为27.43%,以2017年最高;年度蚤指数在2.42~3.39之间,平均蚤指数为2.87,以2012年最高。年度蚤构成比在7.54%~12.15%之间,以2012年最高,见表 1。
捕获的15 602匹鼠体蚤隶属3科7属7种,方叶栉眼蚤(Ctenophthalmus quadratus)占捕获总数的52.10%,为优势蚤种;棕形额蚤指名亚种(Frontopsylla spadix spidix)占28.98%,特新蚤指名亚种(Neopsylla specialis specialis)占17.72%,为常见蚤种;低地狭臀蚤(Stenischia humilis)、缓慢细蚤(Leptopsylla segnis)、无值大锥蚤(Macrostylophora euteles)和不等单蚤(Monophsyllus anisus)分别占0.88%、0.27%、0.04%和0.01%,见表 2。
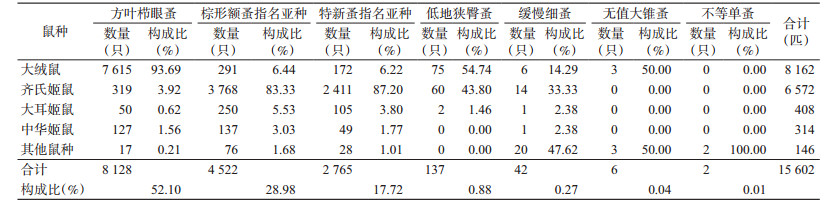
捕获的15 602匹体蚤,分别寄生在2目3科6属11种小型兽类的体表,其中,方叶栉眼蚤的主要寄主是大绒鼠(Eothenomys miletus),棕形额蚤指名亚种的主要寄主是齐氏姬鼠(A. cheurieri),特新蚤指名亚种的主要寄主是齐氏姬鼠,其余4种体蚤数量较少,主要寄主是大绒鼠或齐氏姬鼠,见表 2。
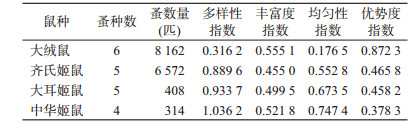
2.4 体蚤的群落多样性特征对体蚤构成比>0.5%的宿主群落进行多样性分析,寄主H在0.316 2~1.036 2之间,中华姬鼠最高;R在0.455 0~0.555 1之间,4种寄主相差不大;J在0.176 5~0.747 4之间,中华姬鼠最高;D在0.378 3~0.872 3之间,大绒鼠最高,见表 3。
蚤类作为鼠疫的传播媒介,染蚤率的高低,与鼠疫在鼠间的流行强度以及波及人间的机会密切相关。2008-2017年的监测中,染蚤率最高年份达到34.30%,对人的潜在威胁加大,建议对该点加强监测,并及时开展预防性的灭鼠、灭蚤工作。
不同蚤种传播鼠疫的能力不同。剑川县野鼠鼠疫疫源地2008-2017年监测结果小型兽类体蚤有7种,方叶栉眼蚤为优势蚤种,棕形额蚤指名亚种和特新蚤指名亚种为常见蚤种,1975-2017年42年间,从剑川县鼠疫疫源地小型兽类体蚤中共分离到43株鼠疫菌[2-3],其中分离自方叶栉眼蚤30株、特新蚤指名亚种7株、棕形额蚤指名亚种4株、低地狭臀蚤和锐额狭臀蚤(Stenischia angustifrontalis)各1株。实验室证实,方叶栉眼蚤对大绒鼠的媒介效能和媒介指数测定均为0,故确认方叶栉眼蚤不具鼠疫媒介作用[6];棕形额蚤指名亚种是云南省野鼠鼠疫疫源地的主要传播媒介之一,也可能是野鼠鼠疫传入家鼠引起人间鼠疫流行的桥梁[7];经实验室证实,特新蚤指名亚种是云南省野鼠鼠疫疫源地的主要传播媒介[8]。此次监测结果,棕形额蚤指名亚种和特新蚤指名亚种为鼠疫的主要传播媒介,方叶栉眼蚤虽不是鼠疫的传播媒介,但该蚤是该鼠疫疫源地的优势蚤种,种群数量较多,构成比达52.10%,而且该蚤会骚扰人、畜的生活,因此,除对特新蚤指名亚种、棕形额蚤指名亚种重点防控外,方叶栉眼蚤也是需要防控的主要蚤类。
影响群落H的因素有丰富度和均匀度,丰富度即物种数,物种数越多,H值越大,各物种间个体分配越均匀,H值就越大[4, 9]。群落中种群数量的分配模式是影响群落J的重要因素[5, 9],单一种类突出的优势是群落J低的主要原因。生态优势度反映群落各物种种群数量的变化情况,群落中物种数量分布越不均匀,其D越大。综合分析剑川县小型兽类体蚤的群落多样性特征,体蚤在大绒鼠和齐氏姬鼠的优势地位突出。
鼠疫主要传播媒介除有很强的传播能力外,还与宿主一起对长期保存鼠疫自然疫源性起重要作用[10-16]。1975-2017年从小型兽类共分离到105株鼠疫菌[2-3],其中分离自大绒鼠39株(占37.14%),齐氏姬鼠46株(占43.81%),大足鼠(Rattus nitidus)11株(占10.48%),大耳姬鼠(A. speciesus)3株(占2.86%),黄胸鼠(R. tanezumi)2株(占1.90%),巢鼠(Micromys minutus)、褐家鼠(R. norvegicus)、北社鼠(Niviventer confucianus)和中华姬鼠各1株,各占0.95%。齐氏姬鼠为该鼠疫疫源地的贮存宿主,大绒鼠是该鼠疫疫源地的主要流行宿主。当小型兽类感染鼠疫菌时,鼠体蚤吸食小型兽类的血液,感染鼠疫菌的概率会增加,感染人的机会也增大。因此,在监测过程中,要密切注意蚤类种群数量和主要宿主的鼠密度,当两者超过预警值时,应及时采取预防性灭鼠、灭蚤措施,及时降低鼠密度和蚤指数,使小型兽类体蚤感染鼠疫菌的机会降低,有效预防鼠疫疫情的发生。
| [1] |
纪树立. 鼠疫流行简史与现状[M]. 北京: 人民卫生出版社, 1986: 2-24.
|
| [2] |
李吉瑞. 剑川鼠疫疫源地监测[J]. 中国地方病学杂志, 2000, 19(6): 482-483. DOI:10.3760/cma.j.issn.1000-4955.2000.06.035 |
| [3] |
蔡文凤, 罗剑龙, 苏丽琼, 等. 云南省剑川县2008-2017年野鼠鼠疫疫源地鼠巢蚤调查[J]. 中国热带医学, 2018, 18(12): 1236-1238. DOI:10.13604/j.cnki.46-1064/r.2018.12.16 |
| [4] |
Margalef DR. Information theory in ecology[J]. Int J Gen Syst, 1958, 3: 36-71. |
| [5] |
Putman RJ, Wratten SD. Principles of ecology[M]. Berkeley: University of California Press, 1984: 320-336.
|
| [6] |
梁云, 何晋侯, 赵文红, 等. 方叶栉眼蚤传播鼠疫媒介效能的实验研究[J]. 地方病通报, 1996, 11(1): 21-23. |
| [7] |
赵文红, 何晋侯, 张洪英, 等. 棕形额蚤传播鼠疫媒介效能的实验研究[J]. 中国地方病防治杂志, 1996, 11(5): 274-275. |
| [8] |
张洪英, 何晋侯, 赵文红, 等. 特新蚤指名亚种传播鼠疫的媒介效能实验研究[J]. 地方病通报, 1996, 11(1): 24-26. DOI:10.13215/j.cnki.jbyfkztb.1996.01.009 |
| [9] |
孙儒泳. 动物生态学原理[M]. 3版. 北京: 北京师范大学出版社, 2017: 399.
|
| [10] |
蔡文凤, 张福新, 王国良, 等. 云南丽江鼠疫疫源地小型兽类鼠体蚤的构成及群落多样性特征[J]. 疾病预防控制通报, 2015, 30(6): 4-7. DOI:10.13215/j.cnki.jbyfkztb.1507013 |
| [11] |
蔡文凤, 张福新, 王国良, 等. 丽江鼠疫自然疫源地犬及猫体蚤调查结果与分析[J]. 中国媒介生物学及控制杂志, 2015, 26(5): 506-508. DOI:10.11853/j.issn.1003.4692.2015.05.020 |
| [12] |
蔡文凤, 张福新, 王国良, 等. 丽江鼠疫自然疫源地地面游离蚤调查[J]. 中国媒介生物学及控制杂志, 2015, 26(6): 617-619. DOI:10.11853/j.issn.1003.4692.2015.06.020 |
| [13] |
程晓藕, 尹家祥. 家鼠鼠疫疫源地蚤类群落结构及其与鼠疫的关系[J]. 中国热带医学, 2015, 15(1): 120-122. |
| [14] |
苏丽琼, 梁云, 吴鹤松, 等. 云南省1986-2014年人间鼠疫流行病学特征分析[J]. 中国媒介生物学及控制杂志, 2016, 27(4): 386-388, 392. DOI:10.11853/j.issn.1003.8280.2016.04.019 |
| [15] |
刘正祥, 高子厚, 尹家祥, 等. 云南省居民区室内小型兽类体表蚤类与宿主关系研究[J]. 中国媒介生物学及控制杂志, 2016, 27(2): 151-154. DOI:10.11853/j.issn.1003.8280.2016.02.015 |
| [16] |
程晓藕, 尹家祥, 王秀芳, 等. 滇西地区室内鼠形动物体表寄生蚤丰盛度影响因素分析[J]. 中华地方病学杂志, 2016, 35(5): 320-324. DOI:10.3760/cma.j.issn.2095-4255.2016.05.003 |