 2018, Vol. 29
2018, Vol. 29扩展功能
文章信息
- 王海峰, 张懿晖, 杨晓燕, 刘溢洋, 牛艳芬, 刘广
- WANG Hai-feng, ZHANG Yi-hui, YANG Xiao-yan, LIU Yi-yang, NIU Yan-fen, LIU Guang
- 两种基因分型方法在河北省鼠疫耶尔森菌基因分型中的应用研究
- Studies on the genotypes of Yersinia pestis in Hebei province by two genotyping methods
- 中国媒介生物学及控制杂志, 2018, 29(6): 564-566, 575
- Chin J Vector Biol & Control, 2018, 29(6): 564-566, 575
- 10.11853/j.issn.1003.8280.2018.06.004
-
文章历史
- 收稿日期: 2018-07-05
- 网络出版时间: 2018-10-16 08:34



河北省鼠疫自然疫源地面积约1 000 km2,自1971年首次判定为鼠疫疫源地至今共发生5次动物间鼠疫流行[1]。河北省鼠疫自然疫源地位于康保县境内,距离首都北京直线距离仅有260 km,距2022年北京冬奥会举办地张家口市崇礼区仅100 km,地理位置特殊,交通便利,旅游业发达,这使得河北省的鼠疫防控工作尤为重要。多位点可变数目串联重复序列(multiple locus variable number tandem repeat analysis,MLVA)和差异区段(different region,DFR)是近年来应用较多,技术成熟的基因分型方法,通过与全国鼠疫耶尔森菌(鼠疫菌)MLVA和DFR基因分型数据比对,可为疫情溯源和流行病学调查提供本底资料。
1 材料与方法 1.1 菌株来源河北省鼠疫疫源地1972—2005年分离的116株鼠疫菌和2017年分离的3株鼠疫菌。
1.2 仪器及试剂PCR扩增仪(杭州博日科技有限公司),高速台式离心机(Eppendorf 5424),水平电泳仪(北京市六一仪器厂),凝胶成像仪(上海基因有限公司)。PCR扩增试剂购自北京全式金生物技术有限公司,DL500 Marker购自大连宝生物工程有限公司,100 bp Marker购自北京赛百盛基因技术有限公司,琼脂糖原产于西班牙Biowest,阳性对照由青海省地方病预防控制所赠送。
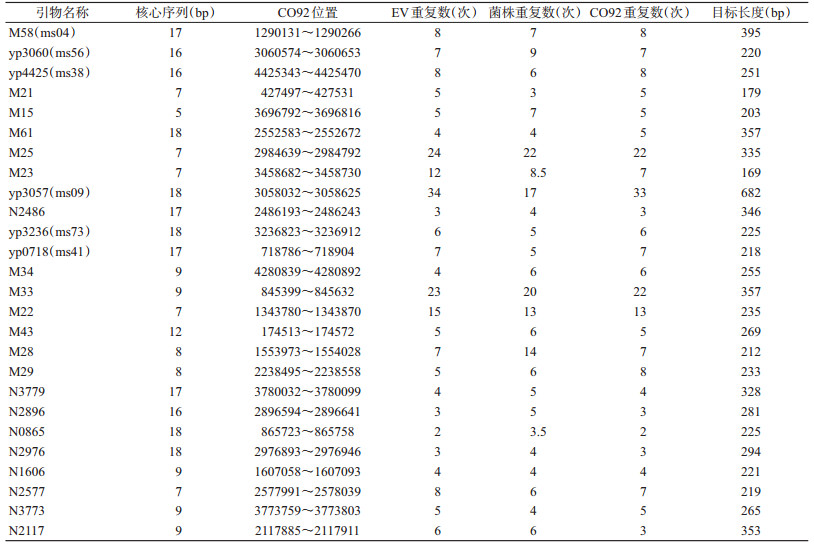
1.3 引物查阅文献,MLVA引物选择“14+12”可变数目串联重复序列(variable-number tandem-repeat,VNTR),DFR引物选择22对DFR和pMT1,由生工生物工程(上海)有限公司合成。引物信息见表 1、2。
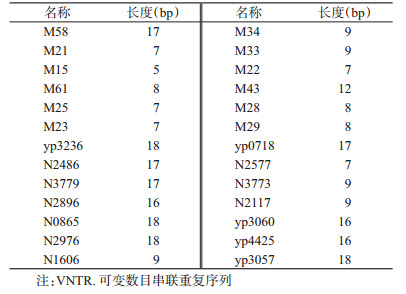
|
用水煮法将分离的鼠疫菌接种于赫氏培养基,28 ℃培养48 h后刮取菌苔于加有600 μl TE缓冲液的Eppendorf管中,100 ℃水浴20 min,离心半径8.4 cm,12 000 r/min离心10 min,收集上清液。抽取100 μl上清液,涂布于赫氏培养基平板中,28 ℃培养48 h,进行无菌实验,将无菌实验阴性的DNA标本用去离子水稀释10倍制备PCR扩增用的模板。
1.4.2 PCR反应体系和条件①反应体系:25 μl反应体系,其中SuperMix混合液12.5 μl,正、反向引物各1.0 μl,模板1.0 μl,补水至25.0 μl;②反应条件:MLVA分型扩增条件为95 ℃预变性5 min,95 ℃变性1 min,引物M58、yp3060、yp4425、M21、M15、M61、M25、M23退火温度为60 ℃,引物yp3057、N2486、yp3236、yp0718、M34、M33、M22、M43、M28、M29退火温度为55 ℃,引物N3779、N2896、N0865、N2976、N1606、N2577、N3773、N2117退火温度为52 ℃,退火时间均为1 min,72 ℃延伸1 min,30个循环;72 ℃延伸5 min;DFR分型引物DFR1~17、DFR20~22、pMT1的扩增条件为94 ℃预变性3 min,94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸1 min。30个循环。72 ℃延伸5 min。引物DFR18和DFR19的退火条件为56 ℃ 30 s,其余条件同DFR1~17、DFR20~22、pMT1。
1.4.3 电泳分析扩增产物用1.5%琼脂糖凝胶电泳,阳性对照采用62004和82009的混合物,阴性对照用去离子水。电泳条件为120V,30~60 min。
1.4.4 结果判读MLVA需要将扩增产物进行测序并统计核心序列重复数目;DFR分型方法中,所有的PCR扩增产物经水平电泳后在凝胶成像仪下观察有无DNA条带,阳性对照扩增出全部条带时结果有效,标本出现目标条带者记为“1”,未出现目标条带者记为“0”,模糊不清的条带重复实验进行验证。
2 结果 2.1 MLVA基因分型结果鼠疫菌26个VNTR[2],每个VNTR位点在116株鼠疫菌中分别表现为相同的重复数,见表 3。
经过多次重复实验,23个DFR中,此阶段分离的116株鼠疫菌扩增产物均缺少DFR01、DFR06、DFR07、DFR13和DFR15~18。通过与文献[3]比对,河北省分离的鼠疫菌归为G17型,见图 1。

|
| 注:1~23.表示引物分别为DFR1~23的鼠疫菌DNA电泳结果 图 1 1972—2005年河北省分离鼠疫菌差异区段分型电泳结果 Figure 1 The electrophoretogram of the wild strain isolated among 1972-2005 amplificated with DFR1-23 |
| |
经过多次重复实验,此次分离的3株鼠疫菌出现目标条带的为DFR02~05、DFR08~11、DFR14、DFR19~22、pMT1;均缺少DFR01、DFR06、DFR07、DFR12、DFR13和DFR15~18。通过与参考文献[4]中鼠疫菌DFR位点的分布和基因型结果比对,确定这3株鼠疫菌的基因型为G20,见图 2。

|
| 注:1~23.表示引物分别为DFR1~23的鼠疫菌DNA电泳结果 图 2 2017年河北省康保县分离鼠疫菌株差异区段分型PCR产物电泳结果 Figure 2 The electrophoretogram of the wild strain isolated in 2017 amplificated with DFR1-23 |
| |
河北省鼠疫疫源地包括长爪沙鼠(Meriones unguiculatus)和达乌尔黄鼠(Spermophilus dauricus)两种类型疫源地,经鉴定分离菌株生态型均属于鄂尔多斯高原型,是内蒙古长爪沙鼠鼠疫疫源地的延伸[5]。但由于地理环境的不同,菌株的遗传特征也不同,所以分离自河北省的鼠疫菌其DFR基因型主要为G17,而内蒙古自治区(内蒙古)主要为G11,G17与G11相比缺失了DFR18片段,基因型的不同导致表型也不同[6],生化实验也证实两地的菌株对蜜二糖和麦芽糖的酵解能力不同[7]。MLVA实验同样证实了此种差别,有的VNTR在两地的菌株中重复数目不同。但是河北省鼠疫疫源地与内蒙古长爪沙鼠鼠疫疫源地自然相连,其间无明显的高山或河流等自然地理屏障,且流行并不频繁,菌株变异机会少,而且流行区域比较集中在与内蒙古正镶白旗、太仆寺旗以及化德县相邻的康保县北部,1972—2005年分离的菌株未发生基因改变,MLVA和DFR分型均只出现1个基因型。
3.2 2017年分离3株菌基因型特征康保县境内在距离上次动物间鼠疫11年的2017年再次发生疫情,流行地区与内蒙古化德县仅约500 m的距离,分离菌株经鉴定DFR基因型为G20,与G17相比缺失了DFR12,与文献报道化德县分离的鼠疫菌基因型一致[8],此次疫情可能是化德县动物间鼠疫疫情的蔓延,而不是河北省鼠疫疫源地内以往流行的鼠疫菌发生了遗传变异,但尚需要进一步收集菌株进行基因分型验证。
3.3 建立基因型型别的意义紧邻的内蒙古长爪沙鼠鼠疫疫源地几乎每年均发生动物疫情,因此鼠疫的输入性风险很大,鼠疫菌的基因型更能反映鼠疫菌的亲缘关系[9],建立准确的鼠疫菌基因型本底资料对于发生疫情及时追根溯源有十分重要的意义,同时也为研究本疫源地菌株的遗传进化规律奠定基础。
| [1] |
杨顺林, 史献明, 杜国义, 等. 河北省动物间鼠疫流行病学分析[J]. 医学动物防制, 2009, 25(4): 247-249. DOI:10.3969/j.issn.1003-6245.2009.04.03 |
| [2] |
Li YJ, Dai EH, Cui YJ, et al. Different region analysis for genotyping Yersinia pestis isolates from China[J]. PLoS One, 2008, 3(5): e2166. DOI:10.1371/journal.pone.0002166 |
| [3] |
周冬生, 韩延平, 宋亚军, 等. 鼠疫耶尔森菌基因组进化与生态位适应研究[J]. 解放军医学杂志, 2004, 29(3): 204-210. DOI:10.3321/j.issn:0577-7402.2004.03.005 |
| [4] |
杨晓艳, 魏柏青, 靳娟, 等. 中国鼠疫耶尔森菌差异区段分型及其地理分布特征[J]. 中华流行病学杂志, 2014, 35(8): 943-948. DOI:10.3760/cma.j.issn.0254-6450.2014.08.015 |
| [5] |
刘合智, 刘满福, 李玉贵. 河北省鼠疫自然疫源地内自然染疫蚤的研究[J]. 中国媒介生物学及控制杂志, 2005, 16(3): 206-208. DOI:10.3969/j.issn.1003-4692.2005.03.017 |
| [6] |
杨瑞馥, 黄培堂. 鼠疫耶尔森菌比较和进化基因组学研究[J]. 解放军医学杂志, 2004, 29(3): 189-191. DOI:10.3321/j.issn:0577-7402.2004.03.001 |
| [7] |
董国润, 王桂琴, 白晓英, 等. 115株鼠疫菌的生物学特性及流行病学研究[J]. 中国媒介生物学及控制杂志, 2001, 12(2): 112-114. DOI:10.3969/j.issn.1003-4692.2001.02.010 |
| [8] |
李建云, 武正华, 胡艳红, 等. 2016年内蒙古长爪沙鼠疫源地化德县鼠疫菌的生态及DFR分型[J]. 医学动物防制, 2017, 33(4): 378-380. |
| [9] |
崔玉军, 宋亚军, 杨瑞馥. 鼠疫耶尔森氏菌的进化研究:从系统发育学到系统发育基因组学[J]. 中国科学:生命科学, 2013, 43(1): 23-30. |