 2017, Vol. 28
2017, Vol. 28扩展功能
文章信息
- 刘蓉蓉, 葛德燕, 鲁亮, 夏琳, 刘起勇, 杨奇森
- LIU Rong-rong, GE De-yan, LU Liang, XIA Lin, LIU Qi-yong, YANG Qi-sen
- 中国姬鼠属种类的DNA条形码鉴定及其分布
- Identification and distribution of Apodemus species with DNA barcoding in China
- 中国媒介生物学及控制杂志, 2017, 28(2): 97-103
- Chin J Vector Biol & Control, 2017, 28(2): 97-103
- 10.11853/j.issn.1003.8280.2017.02.001
-
文章历史
- 收稿日期: 2016-10-31
- 网络出版时间: 2017-01-06 16:46




2 中国科学院动物研究所, 动物进化与系统学院重点实验室, 北京 100101
2 Institute of Zoology, Chinese Academy of Sciences
姬鼠属(Apodemus)种类在我国分布广泛,是我国林地和农田等生境的重要鼠种。其可传播多种人兽共患病,对肾综合征出血热(HFRS)和鼠疫影响较大,尤其是姬鼠型HFRS主要分布在我国中东部,姬鼠传播的鼠疫则局限于滇西北地区,对当地居民威胁相对较小[1-3]。此外,姬鼠属种类在农业、林业、生态等方面均扮演重要角色,是植物保护和生态系统中的重要环节。
我国姬鼠属分类及其系统关系的研究目前还存在许多争议。《中国啮齿类》 [4]涉及6种姬鼠,《中国哺乳动物分布》 [5]记录了6种姬鼠及其分布情况,《Mammal species of the world》 [6]列出分布在我国的有7种,《啮齿动物学》(第2卷)[7]涉及了8种,《中国哺乳动物种和亚种分类名录与分布大全》 [8]收录了9种,而《中国哺乳动物多样性》 [9]中收录了10种,除分布于中国台湾地区的台湾姬鼠(Ap. semotus)外,中国大陆有9种。部分姬鼠属种类的分布情况经长期论证后逐渐清晰,如小林姬鼠(Ap. sylvaticus)在我国的分布情况及澜沧江姬鼠(Ap. ilex)的分类地位[10]。但部分鼠种的分类地位尚存在争议,如蒋学龙和王应祥[11]提出的长尾姬鼠(Ap. orestes)在《啮齿动物学》中并未被认可。因此,对我国姬鼠属鼠种的分类进行深入研究十分迫切。关于姬鼠属种类的分布仍存在较多的不确定性[12]。在许多鼠种同域分布的情况下,只有对某一区域鼠种进行准确鉴定,才能从宏观上汇总各地的鼠种分布情况,进而确定不同鼠种在全国范围内的分布及其界限。针对形态学鉴定鼠种存在的不足,大量文献表明利用DNA条形码技术对鼠种进行鉴定是可行的[13-16]。因此,利用该技术对我国姬鼠属样本进行系统的鉴定,从而确定种类及其分布范围。本研究以采自中国大陆22个省(直辖市、自治区)59个采样点的姬鼠属样本为对象,利用传统的形态学方法及DNA条形码技术对其进行准确鉴定,从而了解其在我国的分布范围及规律。
1 材料与方法 1.1 样本来源研究样本由中国疾病预防控制中心传染病预防控制所媒介生物控制室采自全国各地39个采集点,以及由中国科学院动物研究所采自全国20个自然保护区,从中选取119份代表样本进行研究,样本均保存于-80 ℃。
1.2 形态学鉴定野外捕获到的鼠类,根据其形态学特征进行鉴定并记录,无法在现场鉴别的样本,则制成标本并将其形态相关测量数据带回实验室,根据检索表进行鉴定。
1.3 DNA提取、PCR扩增及测序采用磁珠法对鼠的肝脏或肌肉组织进行DNA提取,置于4 ℃暂存备用。利用通用保守引物COⅠB(5′-CCT ACT CAG GCC ATT TTA CCT ATG-3′)和CO Ⅰ R(5′-ACT TCT GGG TGT CCA AAG AAT CA-3′)扩增细胞色素C氧化酶亚基Ⅰ(COⅠ)基因片段,其反应体系为25 μl,包括Mix(MCLAB)12.5 μl,上下游引物各1 μl(10 μmol/L),DNA模板1.5 μl,ddH2O 9 μl。PCR反应条件为95 ℃预变性5 min,95 ℃变性30 s,50 ℃退火30 s,72 ℃延伸1 min,35个循环,72 ℃延伸3 min。随后用1%琼脂糖凝胶电泳检测扩增产物,并送至北京天一辉远生物科技有限公司进行双向测序。
1.4 序列比对采用Chromas 2软件查看峰图并判断其质量。利用Mega 6.0软件中的MUSCLE对测序获得的有效样本序列进行比对[17],采用邻接法(Neighbor-Joining method,NJ)构建分子系统发育树[18-19]。系统发育树中的褐家鼠(Rattus norvegicus)和大仓鼠(Tscherskia triton)为外群。选择系统发育树中代表个体的序列进行Blast比对[20-21],确定分子鉴定结果,并将GenBank中相似度最高的序列与本研究的序列再次构建系统发育树。
2 结果 2.1 传统形态学鉴定姬鼠属样本经形态学鉴定,包括中华姬鼠(Ap. draco)13只、齐氏姬鼠(Ap. chevrieri)12只、黑线姬鼠(Ap. agrarius)30只、朝鲜姬鼠(Ap. peninsulae)25只、小林姬鼠(Ap. sylvaticus)4只、大耳姬鼠(Ap. latronum)8只及27只待鉴定标本,见表 1。

将Bootstrap Replications值设为1 000。系统发育树显示包括7个姬鼠属鼠种(澜沧江姬鼠、中华姬鼠、大耳姬鼠、朝鲜姬鼠、齐氏姬鼠、乌拉尔姬鼠和黑线姬鼠)及1个待确认种,见图 1。各分支支持率为98%~100%。其中5个分支有对应的GenBank序列,结合形态特点确定鉴定结果,澜沧江姬鼠根据标本的细胞色素b(Cytb)基因确定,而大耳姬鼠仅根据形态学鉴定结果予以确定。

|
| 注:各分支旁的数字为该分支的Bootstrap支持度。 图 1 基于线粒体COⅠ基因构建的姬鼠属种类NJ系统发育树 Figure 1 Molecular phylogeny tree of Apodemus samples based on COⅠ gene with Neighbor-Jioning method |
| |
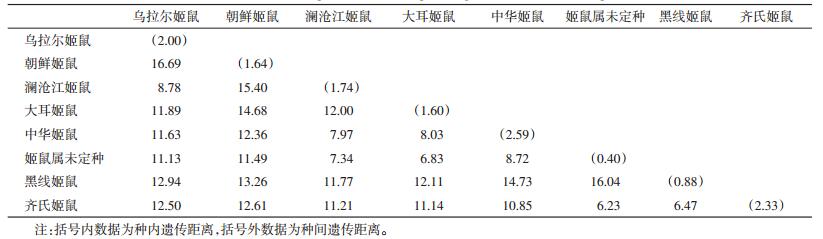
本研究共有8个姬鼠属种类,基于Kimura 2-parameter(K2P)计算其遗传距离。其中中华姬鼠的个体间遗传距离最大,为2.59%;黑线姬鼠虽分布范围广,但遗传距离仅为0.88%;姬鼠未定种个体间遗传距离最小,为0.40%。种间遗传距离为6.23%~16.69%,最大的为乌拉尔姬鼠与朝鲜姬鼠,见表 2。种内和种间遗传距离的变化基本符合其他鼠种种群间的分布。
基于DNA条形码的鉴定结果绘制各鼠种的分布图,见图 2。

|
| 图 2 8种姬鼠属种类在我国的分布情况 Figure 2 The distribution of eight species of Apodemus in China |
| |
根据系统发育树结果,共有21只鼠类样本的分子鉴定结果与形态学鉴定结果有差异(表 1)。总体而言,黑线姬鼠的分子鉴定与形态学鉴定结果一致性最好,但偶尔因鼠类标本皮毛被淋湿等原因可能导致鉴定结果错误;而无典型特征的鼠种间更易出现鉴定错误,如齐氏姬鼠与朝鲜姬鼠、大耳姬鼠与中华姬鼠等。尤其是在2个种均有分布的地区更易出现错误的鉴定结果。
未进行形态学鉴定的鼠类样本中,编号为HB-BD36、LJS009、WL140337和ZT001的鼠种确认为齐氏姬鼠,NM130和NM131为朝鲜姬鼠,WL15447和Sch05050为大耳姬鼠,LJ24、LJ28和JD099为澜沧江姬鼠,QLF14028、WL15097、Sch05054、Sch05086、PQ10、PQ19、QY21及QY31为中华姬鼠(表 1)。采自贵州省江口县梵净山国家级自然保护区的8只样本独立聚为一支,在GenBank中无同源序列,其准确的鉴定结果有待进一步研究。
通过DNA条形码技术对所有鼠类样本进行鉴定后,基本确定我国大陆的姬鼠属种类≥8种,其中澜沧江姬鼠被确定为一独立种,主要分布于云南省横断山脉中南部和西藏自治区东部的高山峡谷地带。从云南省临沧大雪山的采集记录发现,该种主要分布在当地海拔2 500~3 200 m的常绿阔叶林至高山灌丛等生境中,且为该生境的常见种。喜马拉雅姬鼠(Ap. pallipes)分布在西藏自治区的西南区域[9],本研究未在该地区采集标本,故未对该鼠种的分类地位进行讨论。蒋学龙和王应祥[11]提出长尾姬鼠(Ap. orestes)分布于云南省无量山,因本研究无该地区样本,故无法确定。
基于DNA条形码的鉴定结果绘制各鼠种的分布图(图 2),其在省级区域的分布与文献[4, 8]的记载基本一致,说明根据形态特征可区分姬鼠属的大部分鼠种。但与《中国哺乳动物分布》 [5]的分布图有较大差异。对于中华姬鼠与澜沧江姬鼠,仅根据形态特征还无法进行准确区分,原文献也未提及分布信息,本研究可根据分子鉴定结果得出初步的分布信息。本研究认为我国分布最广的姬鼠属种类为黑线姬鼠,除广东、广西、西藏、云南省(自治区)及台湾地区外,其他省(直辖市、自治区)均有分布,尤其是新疆地区西北部阿尔泰山南侧的农耕地区也有分布,且该种主要分布于这些地区的农田及相邻生境中,体现了与人类耕作的密切关系。根据种内遗传距离可以发现,黑线姬鼠虽分布广,但其种内遗传距离较小,说明其存在快速扩散的过程。朝鲜姬鼠分布在与胡焕庸线基本平行但偏西北地区的林地中,仅在东北地区向南扩散到长白山林地,而在中条山(山西省历山)、秦岭南麓(南郑、宁陕县)及横断山脉均未发现该种。齐氏姬鼠主要分布在秦岭以南、大巴山以西的我国第二阶梯区域;而中华姬鼠在陕西和甘肃省一带更向北方扩散到秦岭北侧(宁夏回族自治区六盘山和历山),向东则扩散到华东地区的一些山地高海拔地区(浙江省天目山和百山祖),以及模式产地武夷山,提示该种经历过冰期的南扩过程。在秦岭南坡,齐氏姬鼠的分布稍多于中华姬鼠(南郑和宁陕县)。仅分布于我国西南地区的姬鼠属种类有3种,其中大耳姬鼠虽分布在云南、四川省和西藏自治区,但主要在高海拔地区,在现场调查中,易与中华姬鼠相混淆。澜沧江姬鼠主要分布在横断山脉及以西的西藏地区。而本研究发现的姬鼠属未定种仅分布于贵州省江口县梵净山国家级自然保护区,在贵州省其他采样点未发现。
| [1] | 余向华, 张孝和, 倪庆翔, 等. 鼠传疾病流行病学研究进展[J]. 中国媒介生物学及控制杂志, 2015, 26(6): 634–636. |
| [2] | 王芹, 李建东, 张全福, 等. 2014年全国肾综合征出血热监测总结和疫情分析[J]. 疾病监测, 2016, 31(3): 192–199. |
| [3] | 杨海, 蔡正华, 宗红. 试论姬鼠起源和姬鼠型出血热[J]. 国际病毒学杂志, 2009, 16(3): 94–97. |
| [4] | 黄文几. 中国啮齿类[M]. 上海: 复旦大学出版社, 1995: 122-130. |
| [5] | 张荣祖. 中国哺乳动物分布[M]. 北京: 中国林业出版社, 1997: 188-193. |
| [6] | Wilson DE, Reeder DM. Mammal species of the world:a taxonomic and geographic reference[M]. 3rd ed. Baltimore: The Johns Hopkins University Press, 2005: 1259-1280. |
| [7] | 郑智民, 姜志宽, 陈安国. 啮齿动物学.第2卷[M]. 2版. 上海: 上海交通大学出版社, 2012: 57-74. |
| [8] | 王应祥. 中国哺乳动物种和亚种分类名录与分布大全[M]. 北京: 中国林业出版社, 2003: 193-196. |
| [9] | 蒋志刚, 马勇, 吴毅, 等. 中国哺乳动物多样性[J]. 生物多样性, 2015, 23(3): 351–364. |
| [10] | Liu XM, Wei FW, Li M, et al. Molecular phylogeny and taxonomy of wood mice (genus Apodemus Kaup, 1829) based on complete mtDNA cytochrome b sequences, with emphasis on Chinese species[J]. Mol Phylogenet Evol, 2004, 33(1) : 1–15 .DOI:10.1016/j.ympev.2004.05.011. |
| [11] | 蒋学龙, 王应祥. 长尾姬鼠分类地位的探讨[J]. 动物学研究, 2000, 21(6): 473–478. |
| [12] | 杨海, 王戬. 姬鼠系统分类研究[J]. 中华地方病学杂志, 2008, 27(5): 587–589. |
| [13] | 何锴, 王文智, 李权, 等. DNA条形码技术在小型兽类鉴定中的探索:以甘肃莲花山为例[J]. 生物多样性, 2013, 21(2): 197–205. |
| [14] | Li J, Zheng X, Cai YS, et al. DNA barcoding of Murinae (Rodentia:Muridae) and arvicolinae (Rodentia:Cricetidae) distributed in China[J]. Mol Ecol Res, 2014, 15(1) : 153–167 . |
| [15] | Ondrejicka DA, Locke SA, Morey K, et al. Status and prospects of DNA barcoding in medically important parasites and vectors[J]. Trends Parasitol, 2014, 30(12) : 582–591 .DOI:10.1016/j.pt.2014.09.003. |
| [16] | 金圣浩, 鲁亮, 闫东, 等. DNA条形码技术在河北省坝上地区夜行鼠鉴定中的应用[J]. 中国媒介生物学及控制杂志, 2015, 26(1): 55–57. |
| [17] | Tamura K, Stecher G, Peterson D, et al. Mega 6:molecular evolutionary genetics analysis version 6.0[J]. Mol Biol Evol, 2013, 30(12) : 2725–2729 .DOI:10.1093/molbev/mst197. |
| [18] | Atteson K. The performance of Neighbor-Joining methods of phylogenetic reconstruction[J]. Algorithmica, 1999, 25(2/3) : 251–278 . |
| [19] | Gascuel O, Steel M. Neighbor-Joining revealed[J]. Mol Biol Evol, 2006, 23(11) : 1997–2000 .DOI:10.1093/molbev/msl072. |
| [20] | Altschul SF, Madden TL, Schäffer AA, et al. Gapped BLAST and PSI-BLAST:a new generation of protein database search programs[J]. Nucleic Acids Res, 1997, 25(17) : 3389–3402 .DOI:10.1093/nar/25.17.3389. |
| [21] | Lobo I. Basic local alignment search tool (BLAST)[J]. J Mol Biol, 1990, 215(3) : 403–410 .DOI:10.1016/S0022-2836(05)80360-2. |