 2017, Vol. 28
2017, Vol. 28扩展功能
文章信息
- 韩晓莉, 黄钢, 王喜明, 王岩, 马丽华, 高文
- HAN Xiao-li, HUANG Gang, WANG Xi-ming, WANG Yan, MA Li-hua, GAO Wen
- 河北省德国小蠊野外种群抗药性水平与酶活性关系的通径分析
- Path analysis of the relationship between the resistance level and enzyme activity of field population of Blattella germanica in Hebei province
- 中国媒介生物学及控制杂志, 2017, 28(6): 567-571
- Chin J Vector Biol & Control, 2017, 28(6): 567-571
- 10.11853/j.issn.1003.8280.2017.06.013
-
文章历史
- 收稿日期: 2017-06-20
- 网络出版时间: 2017-08-09 16:08




由于杀虫剂的广泛和频繁使用而导致的害虫抗药性发生、发展,使德国小蠊(Blattella germanica)已成为全球最普遍、最难治理的城市卫生害虫[1],国内很多省份均逐步开展了抗性监测工作。据报道,有50%左右的地区其抗性已达到中抗及其以上水平,且其抗性水平呈逐渐上升趋势[2]。河北省近年的抗药性监测结果也表明,河北省各地区德国小蠊野外种群对常用杀虫剂均已产生不同程度的抗性[3],抗性增强导致杀虫剂使用效率下降,加大了防治难度。对德国小蠊抗药性机制的研究有助于抗药性治理和制定有效的防制策略,延缓抗性水平的上升。目前普遍认为代谢抗性和靶标抗性是害虫抗药性产生的最主要和最普遍机制[4],乙酰胆碱酯酶(AChE)、谷胱甘肽-S-转移酶(GSTs)、羧酸酯酶(CarE)和多功能氧化酶(MFO)是德国小蠊体内重要的靶标酶和代谢酶,德国小蠊抗药性的形成与体内各种酶系的活性变化存在密切联系,国内对AChE、CarE和GSTs的研究报道较多[5-10],王学军[8]研究发现德国小蠊野外种群在AChE和GSTs活性上存在差异,吴崧霖等[10]研究结果显示,GSTs在野外种群的抗性形成过程中起一定作用,而MFO活力增强是导致德国小蠊抗药性产生和发展的重要原因[11],作为与昆虫抗性水平相关的这几种酶,其活性大小也会影响不同杀虫剂的抗性水平。本研究在测定河北省不同地区德国小蠊野外种群AChE、GSTs、CarE、MFO活性和抗性水平的基础上,用通径分析法探讨这几种酶的活性与抗性水平的关系以及其在德国小蠊对不同杀虫剂的抗药性中的作用,旨在为德国小蠊防制中合理使用杀虫剂及抗性治理提供依据。
1 材料与方法 1.1 试虫来源德国小蠊成虫和若虫采自承德、张家口、唐山、保定、沧州、邢台、邯郸和秦皇岛8个市,经饲养室饲养1~2代后,取2~3周龄成虫供试。对照组试虫为河北省CDC媒介昆虫饲养室常年饲养,不接触任何杀虫剂。
1.2 主要化学试剂及仪器主要试剂有碘化硫代乙酰胆碱、α-萘酚、牛血清白蛋白、对硝基苯酚、α-乙酸萘酯,国药集团化学试剂有限公司;还原型谷胱甘肽(GSH)、还原型辅酶Ⅱ(NADPH)、二硫双对硝基苯甲酸(DTNB)、考马斯亮蓝G-250,Solarbio;1-氯-2,4-二硝基苯(CDNB)、毒扁碱、坚固蓝B盐,进口分装;对硝基苯甲醚,东京化成工业株式会社;乙二胺四乙酸,天津永大化学试剂有限公司。主要仪器:耶拿specord 200型紫外分光光度计,sigma 1-14K超高速冷冻离心机,博讯SHZ-A水浴振荡器。
1.3 方法 1.3.1 酶源制备 1.3.1.1 CarE挑取20只德国小蠊成虫,放入玻璃匀浆器中,加入2 ml 0.04 mol/L pH 7.0磷酸盐缓冲液(PBS),在冰浴条件下匀浆,加入1 ml PBS稀释,在4 ℃,离心半径6.6 cm,10 000 r/min离心10 min,取上清液,加入3 ml 0.04 mol/L pH 7.0 PBS混匀,冰浴待用。
1.3.1.2 AChE取20只德国小蠊成虫,加入3 ml 0.1 mol/L pH 7.5 PBS,放于匀浆器内,在冰浴条件下匀浆,并在4 ℃下,离心半径6.6 cm,10 000 r/min离心5 min,取上清液,加入2 ml 0.1 mol/L pH 7.5 PBS,混匀,冰浴待用。
1.3.1.3 MFO挑取20只德国小蠊成虫,放入玻璃匀浆器中,加入1.5 ml PBS(0.2 mol/L,pH 7.8)中充分匀浆,加入1.5 ml PBS稀释,在4 ℃,离心半径6.6 cm,10 000 r/min离心15 min,取上清液做酶液。
1.3.1.4 GSTs挑取20只德国小蠊成虫,放入玻璃匀浆器中,加入2 ml0.2 mol/L pH 6.5 PBS,在冰浴条件下匀浆,加入1 ml PBS稀释,并在4 ℃,离心半径6.6 cm,10 000 r/min离心15min,取上清液,加入1 ml 0.1 mol/L pH 6.5 PBS,混匀,冰浴待用。
1.3.2 活性测定 1.3.2.1 CarE活性测定参照van Asperen[12]的方法并加以修改。在每个试管中依次加入1 ml含有3×10-4 mol/L底物和毒扁豆碱的PBS(0.04 mol/L,pH 7.0),1 ml酶液。30 ℃反应10 min后,加入坚固蓝B缓冲液(1%坚固蓝B盐:5%SDS,2:5显色剂)终止反应并显色,静置15 min后在600 nm下测A值。对照管中在加入坚固蓝B缓冲液之后补加酶液,用α-萘酚制作标准曲线,酶原经蛋白质含量测定,计算CarE比活力〔α-NA/(mg pro·min)〕。
1.3.2.2 AChE活性测定采用Gorun等[13]改进的Ellman方法[14]。取待测酶液0.1 ml与0.1 ml硫代乙酰胆碱(ATCh)混匀,在30 ℃下水浴反应15 min,加入3.6 ml显色剂(DTNB)终止反应,在412 nm处测A值,并计算比活力〔mmol/(mg pro·min)〕。
1.3.2.3 MFO-O-脱甲基活性测定参照Hung和Sun[15]方法并加以改进。反应体系中含0.9 ml 0.2 mol/L pH 7.8 PBS,0.1 ml 0.1 mol/L对硝基苯甲醚,1 ml 1 mol/L NADPH,1 ml酶液。摇匀置于37 ℃下水浴振荡30 min,加入1 ml 1 mol/L HCl终止反应,用5 ml氯仿萃取,弃去水相,再用3 ml 0.5 mol/L NaOH萃取,取水相于400 nm处测A值。对照以缓冲液取代酶液,根据对硝基苯酚标准曲线和酶源蛋白质含量,将A值换算成比活力〔μmol/(mg pro·30 min)〕。
1.3.2.4 GSTs活性测定参照Habig和Willian[16]方法,取2.7 ml0.1 mol/L pH 6.5 PBS,0.1 ml 20 mmol/L GSH,0.1 ml酶液加入到比色杯中,用分光光度计在340 nm下调零,然后加入0.1 ml CDNB快速摇匀,340 nm下测A值,酶原经蛋白质含量测定,计算出比活力〔A/(mg pro·min)〕。
1.4 抗性判定标准敏感群体(S)、抗性群体(R),R/S≤1无抗性;1<R/S≤5为低度抗性;5<R/S≤10为中度抗性;10<R/S≤50为高度抗性;R/S>50为极高抗性。
1.5 统计学处理使用SPSS 19.0软件对试验数据进行分析、处理。采用LSD法对德国小蠊野外种群CarE、AChE、MFO及GSTs的活力进行比较;利用Pearson相关分析酶活性比值与抗药性倍数的相关性;用SPSS线性回归实现酶活性比值与抗药性指数的通径分析。
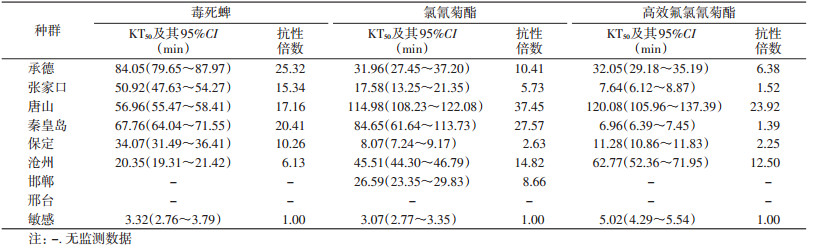
2 结果 2.1 不同地区德国小蠊野外种群对3种杀虫剂的抗性水平不同地区德国小蠊野外种群对毒死蜱、氯氰菊酯和高效氟氯氰菊酯均表现出不同程度的抗性,其中承德种群对毒死蜱的抗性水平最高,抗性倍数为25.32倍,为高抗水平;唐山种群对氯氰菊酯及高效氟氯氰菊酯的抗性水平最高,抗性倍数分别为37.45和23.92倍,均为高抗水平;沧州种群对毒死蜱的抗性水平最低,抗性倍数为6.13倍,为中抗水平;保定种群对氯氰菊酯的抗性水平最低,抗性倍数为2.63倍,为低抗水平;秦皇岛种群对高效氟氯氰菊酯的抗性水平最低,抗性倍数为1.39倍,为低抗水平,见表 1。
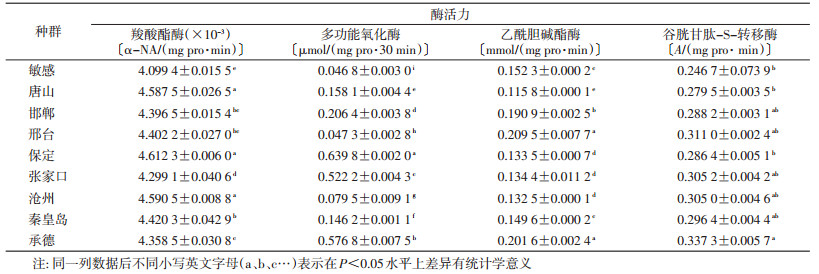
各地德国小蠊野外种群中CarE、MFO-氧-脱甲基活性均明显高于敏感种群,其中CarE的活性以保定种群最高,张家口种群最低;MFO-氧-脱甲基活性以保定种群最高,邢台种群最低;各种群中AChE及GSTs活性差异较大,AChE活性测定结果表明,秦皇岛种群与敏感种群无明显差异,邯郸、承德、邢台种群明显高于敏感种群,张家口、保定、沧州、唐山种群明显低于敏感种群,而GSTs活性测定结果表明,保定、唐山种群与敏感种群无差异,邢台、张家口、沧州、秦皇岛、邯郸、承德种群明显高于敏感种群,见表 2。
 |
相关性分析结果显示,德国小蠊体内MFO与对氯氰菊酯的抗性水平呈一定的相关性,相关系数为-0.767;CarE及AChE活性与对毒死蜱的抗性水平呈一定的相关性,相关系数分别为-0.626和0.687;GSTs与3种杀虫剂抗性水平相关性较小;4种酶活性与不同杀虫剂的抗性水平相关性均未达到显著水平,见表 3。
 |
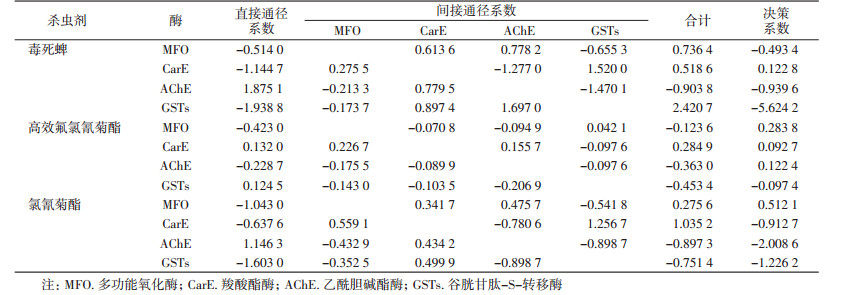
从直接作用来看,直接通径系数越高表明某一酶活性对抗性水平的直接作用越大,不同酶活性对毒死蜱的抗性水平的直接作用表现为AChE>MFO>CarE>GSTs,不同酶活性对高效氟氯氰菊酯的抗性水平的直接作用表现为CarE>GSTs>AChE>MFO,不同酶活性对氯氰菊酯的抗性水平的直接作用表现为AChE>CarE>MFO>GSTs。从间接作用来看,间接通径系数越高表明某一酶活性对抗性水平的间接作用越大,不同酶活性对毒死蜱抗性水平的间接作用为GSTs>MFO>CarE>AChE,不同酶活性对高效氟氯氰菊酯抗性水平的间接作用为CarE>MFO>AChE>GSTs,不同酶活性对氯氰菊酯抗性水平的间接作用为CarE>MFO>GSTs>AChE。决策系数表明某一酶活性对抗性水平的综合作用,决策系数>0表明酶活性对抗性水平起增进作用,决策系数<0表明酶活性对抗性水平起抑制作用,从酶活性对杀虫剂抗药性水平的综合作用来看,CarE活性对毒死蜱抗药性水平起主要增进作用,决策系数为0.122 8;MFO活性对高效氟氯氰菊酯和氯氰菊酯的抗药性水平起主要增进作用,决策系数分别为0.283 8和0.512 1,见表 4。
 |
通径分析以其能区分自变量对因变量的直接作用和间接作用及可根据通径系数大小判断各自变量对因变量贡献的相对重要性等特点,在农业、生物学及流行病学等诸多领域得到广泛应用[17-19]。基于通径分析的这些特点,可以在研究酶活性与抗药性水平的关系时,用以说明酶活性在抗性水平上所发挥的作用,较之以往单独以相关系数来说明二者之间的关系更为全面和直接。本研究结合通径分析从酶活性对抗性水平的直接作用、间接作用及综合作用3个方面来说明酶活性在德国小蠊对不同杀虫剂抗性中的作用,进一步探讨河北省不同地区德国小蠊野外种群的可能抗性机制,为抗性治理和实施有效的化学防治提供依据。
酶活性与抗药性水平的相关性分析结果表明,德国小蠊体内MFO与对氯氰菊酯的抗性水平及CarE活性与对毒死蜱的抗性水平相关系数均出现了负值,且各相关系数差异均无统计学意义,因此仅用相关性分析并不能很好地说明酶活性与抗药性水平的关系,而通径分析法可以弥补这些不足,可利用直接通径系数、间接通径系数和决策系数来说明酶活性在抗性水平中的不同作用。在通径分析结果中,直接通径系数表明某一酶活性对抗性水平起直接作用,间接通径系数表明某一酶活性对抗性水平的影响是通过其他酶的活性影响而起间接作用,决策系数则表明某一酶活性对抗性水平的综合作用,决策系数>0表明酶活性对抗性水平起增进作用,决策系数<0表明酶活性对抗性水平起抑制作用。
各种酶在不同类型的抗药性发展过程中的作用不尽相同,本研究结合相关性分析及酶活性与抗药性水平的通径分析结果显示,CarE在河北省不同地区德国小蠊野外种群对毒死蜱的抗性中起主要增进作用,在对高效氟氯氰菊酯的抗性中起到了直接和间接作用;AChE在德国小蠊对毒死蜱和氯氰菊酯的抗性中起到了直接作用;MFO在德国小蠊对氯氰菊酯和高效氟氯氰菊酯的抗性中起到了主要增进作用;GSTs在德国小蠊野外种群对毒死蜱、氯氰菊酯和高效氟氯氰菊酯3种杀虫剂的抗性水平中有较强的抑制作用,说明GSTs本身并不直接影响抗性水平,但可抑制其他酶对抗性水平的影响。因此表明CarE、AChE和MFO活力变化与德国小蠊对3种杀虫剂的抗药性密切相关。
AChE是有机磷和氨基甲酸酯类杀虫剂的主要作用靶标,AChE敏感性高低与昆虫对有机磷和氨基甲酸酯类杀虫剂的抗性有着密切关系,但本研究发现,并不是全部德国小蠊种群中AChE活性均低于敏感种群,而是表现出与敏感种群比较有高有低,这与其他地区的研究结果类似[10],可能与德国小蠊对作用机制不同的杀虫剂同时表现出抗性有关。在研究中也发现,CarE在德国小蠊野外种群对毒死蜱的抗性中也起主要增进作用,因此就河北省德国小蠊野外种群对毒死蜱的抗性而言,可能与AChE和CarE的活力变化有关。
在各地德国小蠊野外种群对氯氰菊酯的抗性中,酶活性测定结果显示,各野外种群MFO-氧-脱甲基活性明显高于敏感种群,通径分析结果表明MFO起主要增进作用,AChE起直接作用,说明MFO活性的提高是河北省各地德国小蠊野外种群对氯氰菊酯产生抗性的主要原因,AChE也起到了重要作用。
在各地德国小蠊野外种群对高效氟氯氰菊酯的抗性中,由于各野外种群MFO-氧-脱甲基活性及CarE活性明显高于敏感种群,通径分析结果也表明CarE和MFO分别起直接、间接作用和主要增进作用,说明河北省德国小蠊野外种群对高效氟氯氰菊酯的抗性可能与MFO和CarE活力增强有关。
GSTs与3种杀虫剂抗性水平相关性较小,而在通径分析结果中,GSTs在德国小蠊野外种群中对3种杀虫剂的抗性水平有较强的抑制作用,可能是由于GSTs对抗性水平的作用很弱且本身并不直接影响抗性水平,因此,GSTs在河北省德国小蠊野外种群对3种杀虫剂抗药性水平中的作用不大。
对于德国小蠊野外种群而言,由于杀虫剂的使用情况比较复杂,害虫通常存在多抗性,由此导致了德国小蠊野外种群抗性机制的复杂性,本研究从实际需求出发,以相关系数和通径系数相结合,从生化检测的角度分析不同杀虫剂抗性与酶活力的关系,分析了各种酶在德国小蠊对不同杀虫剂抗性水平中的作用,探讨了河北省德国小蠊野外种群的可能抗性机制,为通径分析法在酶活性与抗药性水平中的适用性进行了探索,可为今后抗性机制的研究提供新思路。
| [1] |
霍新北, 刘起勇. 德国小蠊抗药性的种群特性及治理策略[J]. 中国媒介生物学及控制杂志, 2013, 24(2): 89-91. |
| [2] |
王学军, 赖世宏, 赵志刚, 等. 山东省德国小蠊对5种杀虫剂的抗药性调查[J]. 中华卫生杀虫药械, 2012, 18(4): 332-335. |
| [3] |
韩晓莉, 高文, 黄钢, 等. 运用聚类分析法进行德国小蠊抗药性水平区域划分[J]. 中国媒介生物学及控制杂志, 2016, 27(5): 467-469, 473. DOI:10.11853/j.issn.1003.8280.2016.05.011 |
| [4] |
张凡. 德国小蠊抗药性机理[J]. 生命科学, 2011, 31(5): 747-750. |
| [5] |
王学军, 康殿民, 张凡, 等. 德国小蠊抗药性与乙酰胆碱酯酶、谷胱甘肽S-转移酶和超氧化物歧化酶活性关系的研究[J]. 中国媒介生物学及控制杂志, 2013, 24(2): 135-137. |
| [6] |
靖博斌, 刘小闪, 冯丽娟, 等. 高效氯氰菊酯对德国小蠊酯酶活性的影响[J]. 铁路节能环保与安全卫生, 2016, 6(1): 26-29. |
| [7] |
王式春, 林琳, 刘渠, 等. 抗药性与敏感性德国小蠊乙酰胆碱酯酶的活性比较[J]. 热带医学杂志, 2011, 11(7): 748-749. |
| [8] |
王学军. 山东省德国小蠊抗药性监测及抗性相关酶生化特征研究[D]. 济南: 山东大学, 2012. http://d.wanfangdata.com.cn/Thesis/Y2180851
|
| [9] |
曹官时, 霍新北. 溴氰菊酯对德国小蠊AChE及GSTs作用研究[J]. 医学动物防制, 2012, 28(4): 401-403. |
| [10] |
吴崧霖, 吴能简, 陈伟文, 等. 坪山新区德国小蠊谷胱甘肽S-转移酶活性研究[J]. 中国卫生工程学, 2016, 15(5): 429-430. |
| [11] |
张大羽, 程家安, 吴慧明, 等. 德国小蠊的抗药性及多功能氧化酶在抗性中的作用[J]. 浙江农业大学学报, 1999, 25(3): 303-306. |
| [12] |
van Asperen K. A study of housefly esterases by means of a sensitive colorimetric method[J]. J Insect Physiol, 1962, 8(4): 401-416. DOI:10.1016/0022-1910(62)90074-4 |
| [13] |
Gorun V, Porinov L, Bãlteseu V, et al. Modified Ellman procedure for assay of cholinesterases in crude enzymatic preparations[J]. Anal Biochem, 1978, 86(1): 324-326. DOI:10.1016/0003-2697(78)90350-0 |
| [14] |
高希武. Gorun等改进的Ellman胆碱酯酶活性测定方法介绍[J]. 昆虫知识, 1987(4): 245-246. |
| [15] |
Hung CF, Sun CN. Microsomal monooxygenases in diamondback mothlarvae resistant to fenvalerate and piperonylbutoxide[J]. Pestic Biochem Physiol, 1989, 33(2): 168-175. DOI:10.1016/0048-3575(89)90007-2 |
| [16] |
Habig WH, Willian BJ. Method in Enzynology[M]. New York: Academic Press, 1981, 398-405.
|
| [17] |
叶小华, 郜艳晖, 张敏, 等. 高中生健康行为影响因素的多组通径分析[J]. 现代预防医学, 2013, 40(12): 2192-2195. |
| [18] |
卢晓萍, 杨丙贤, 徐婵娟, 等. 3种小筆科植物叶片SPAD值与叶绿素的相关性及通径分析[J]. 浙江大学学报:农业与生命科学版, 2013, 39(3): 261-266. |
| [19] |
武路广, 霍梅俊, 刘思奇, 等. 白羊草干草产量与主要农艺性状的多元回归及通径分析[J]. 草地学报, 2013, 21(4): 697-701. DOI:10.11733/j.issn.1007-0435.2013.04.011 |