 2017, Vol. 28
2017, Vol. 28扩展功能
文章信息
- 汤丽, 张长国, 邵宗体, 段兴德, 浦恩念, 刘正祥, 苏超, 宋志忠, 高子厚
- TANG Li, ZHANG Chang-guo, SHAO Zong-ti, DUAN Xing-de, PU En-nian, LIU Zheng-xiang, SU Chao, SONG Zhi-zhong, GAO Zi-hou
- 云南省玉龙鼠疫疫源地蚤类群落结构与物种多样性
- The community structures and species diversity of fleas in Yulong plague focus, Yunnan province
- 中国媒介生物学及控制杂志, 2017, 28(3): 201-204, 215
- Chin J Vector Biol & Control, 2017, 28(3): 201-204, 215
- 10.11853/j.issn.1003.8280.2017.03.002
-
文章历史
- 收稿日期: 2016-12-29
- 网络出版时间: 2017-04-11 16:05




2 云南省地方病防治所, 云南 大理 671000
2 Provincial Key Laboratory for Zoonosis Control and Prevention, Yunnan Institute of Endemic Disease Control and Prevention
云南省丽江市玉龙鼠疫自然疫源地位于滇西北山区,是2006年被证实和新发现的鼠疫疫源地。一些学者对该疫源地宿主动物和媒介生物中分离的鼠疫耶尔森菌(鼠疫菌)的生化特性和遗传特征等进行了初步研究,基本查清了疫源地的范围[1-6]。但对于该疫源地的宿主动物和媒介生物群落结构特征、物种多样性及多度分布格局等尚未进行系统研究,尤其是对不同海拔高度的蚤类群落组织水平还需进行系统地研究。因此,于2015年3-12月根据不同生境和海拔定期对玉龙鼠疫疫源地小兽及体表寄生蚤类进行调查,了解玉龙鼠疫疫源地蚤类多样性及垂直分布特征,为鼠疫的防治和监测提供基础资料和理论依据。
1 材料与方法 1.1 调查范围和时间根据玉龙鼠疫疫源地植被分布特征,将调查地区划分为4个不同梯度的垂直带和两种生境(林地和耕地),其中Ⅰ带(2 392~2 650 m)为云南松中幼林、灌木丛和耕地,Ⅱ带(2 650~2 800 m)为多变石砾硬阔叶林,Ⅲ带(2 800~3 100 m)为华山松和响叶杨混交林,Ⅳ带(>3 100 m)为光叶石砾、光叶高山栎、稀疏灌木林和石质山地。分别于2015年3-4月、6-7月、8-9月和11-12月对玉龙鼠疫疫源地进行小兽及体表寄生蚤的调查。
1.2 调查方法每次按不同植被带和不同生境(林地和耕地)选取8个采样点进行调查。依据小兽的活动习性,采用夹夜法[7]布放,根据每个采样点地形,先选择4~5条样线,在每条样线上布放鼠夹40~60夹,夹距5 m,样线间距20~50 m,每个采样点的鼠夹≥250夹,以新鲜炸油条为诱饵,晚放晨收。捕获的小兽用小布袋分装,带回实验室麻醉后收集其体表全部蚤类,置70%乙醇保存,依据文献[8]进行计数、分类鉴定。
1.3 数据分析 1.3.1 计算公式构成比(Dr)[9]:Dr=Ni/N;式中Ni为某种蚤类或某种宿主个体数,N为蚤类或宿主总个体数。
1.3.2 群落水平指标测定物种丰富度指数用物种数S表示。优势度指数采用Simpson公式:C′=∑(ni/N)2,式中C′为生态优势度,ni为每一种的个体数,N为总个体数。多样性指数用Shannon-Winner指数公式: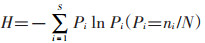
根据小兽及其体表寄生蚤群落中各物种的捕获比例,按照徐正会等[11]提出的依据物种个体数量占样本个体总数的比例,将构成比>10%的种类定为优势种,1%~10%为常见种,<l%为稀有种。
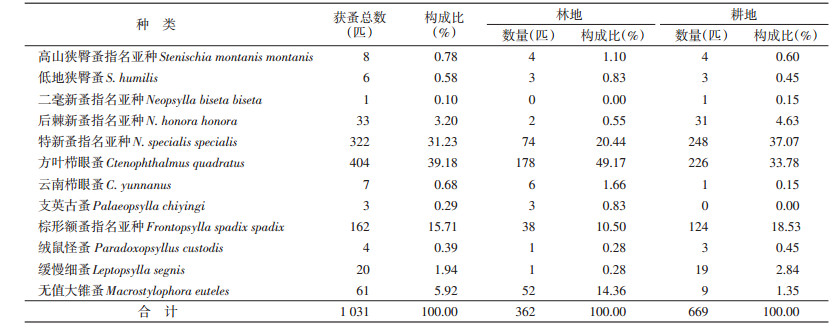
2 结果 2.1 蚤类的数量特征共捕获小兽4目6科12属22种1 575只,检获小兽体表寄生蚤3科8属12种1 031匹。耕地捕获蚤类数量高于林地,分别为669和362匹。两种生境的优势种为特新蚤指名亚种(Neopsylla specialis specialis)、方叶栉眼蚤(Ctenophthalmus quadratus)和棕形额蚤指名亚种(Frontopsylla spadix spadix),占全部蚤类的86.13%(888/1 031);其中林地的优势蚤种为方叶栉眼蚤、特新蚤指名亚种、无值大锥蚤(Macrostylophora euteles)和棕形额蚤指名亚种;耕地的优势种为特新蚤指名亚种、方叶栉眼蚤和棕形额蚤指名亚种,见表 1。
|
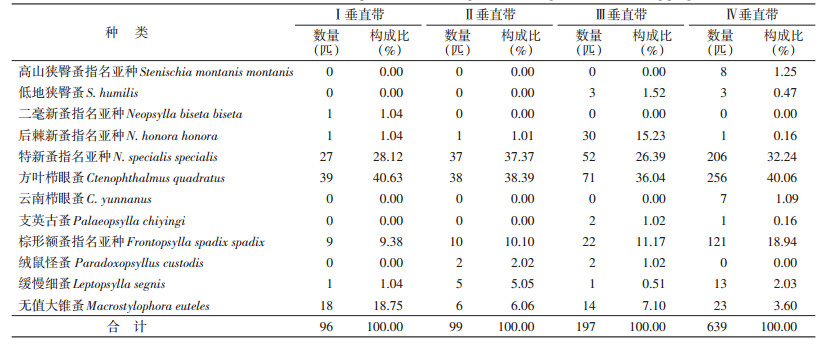
玉龙鼠疫疫源地自然环境以特新蚤指名亚种、方叶栉眼蚤、棕形额蚤指名亚种、缓慢细蚤和无值大锥蚤的垂直分布范围较广,在4个垂直带均有分布,显示这些蚤种对不同植被环境具有较强的适应和占领能力,其余蚤类的分布范围相对要局限一些,部分蚤类垂直分布范围较小,仅局限于1个分布带,如高山狭臀蚤指名亚种、二毫新蚤指名亚种等。疫源地蚤类的分布与海拔密切相关,Ⅰ和Ⅱ垂直带均有7种蚤,以特新蚤指名亚种、方叶栉眼蚤、棕形额蚤指名亚种和无值大锥蚤为主,分别占该区检获蚤类的96.88%(93/96)和91.92%(91/99);Ⅲ垂直带的寄生蚤增加至9种,优势种为特新蚤指名亚种、方叶栉眼蚤、后棘新蚤指名亚种和棕形额蚤指名亚种,占该区检获蚤类的88.83%(175/197);Ⅳ垂直带蚤种丰富度和捕获数量均较高,分别为10种639匹,超过前3个垂直带捕获蚤总数,优势蚤种为特新蚤指名亚种、方叶栉眼蚤和棕形额蚤指名亚种,疫源地蚤类的物种丰富度和蚤数量特征呈现随海拔增加逐步增加的分布趋势,见表 2。
|
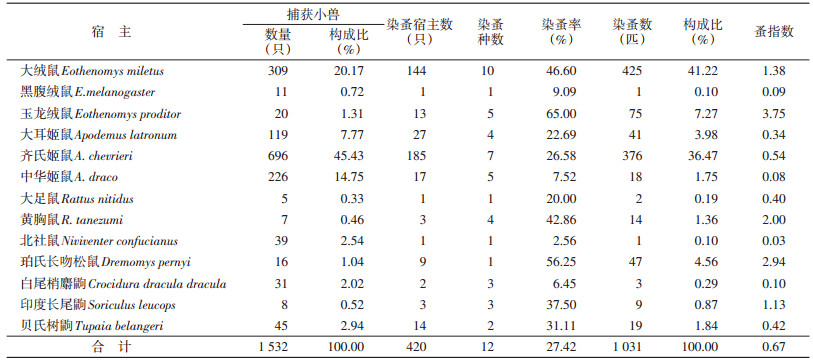
捕获小兽22种,13种染有蚤类。大绒鼠(Eothenomys miletus)染带蚤种最多,共10种,其次是齐氏姬鼠(Apodemus chevrieri)和中华姬鼠(Ap. draco),分别染带7种和5种蚤类,黑腹绒鼠(E. melanogaster)、大足鼠(Rattus nitidus)和北社鼠(Niviventer confucianus)仅染带1种蚤类。检获蚤类绝对数量以方叶栉眼蚤最多,其次是特新蚤指名亚种和棕形额蚤指名亚种。前者主要寄生于大绒鼠,是该鼠寄生蚤中的优势种;后者是齐氏姬鼠寄生蚤中的优势种;而其他蚤类可能是对应宿主动物的常见种或偶然染带。
小兽的染蚤率为27.42%,其中玉龙绒鼠(E. proditor)染蚤率最高,为65.00%;其次是珀氏长吻松鼠(Dremomys pernyi)、大绒鼠、黄胸鼠(R. tanezumi )和齐氏姬鼠,染蚤率分别为56.25%、46.60%、42.86%和26.58%;黑腹绒鼠、中华姬鼠、白尾梢麝鼩和北社鼠染蚤率较低。总蚤指数为0.67,玉龙绒鼠蚤指数最高(3.75),北社鼠最低(0.03),其他小兽蚤指数在0.08~2.94之间,见表 3。
|
玉龙鼠疫疫源地小兽寄生蚤类的物种丰富度随海拔增加呈逐渐增高趋势,在Ⅳ垂直带形成高峰,为10种;蚤类生态优势度以Ⅱ和Ⅳ垂直带略高,分别为0.303 9和0.302 3,优势种方叶栉眼蚤和特新蚤指名亚种的地位突出,相反均匀度指数较低,为0.726 1和0.605 9;而Ⅲ垂直带生态优势度最小,为0.240 7,均匀度较高,为0.738 4;蚤类多样性指数总体趋势呈现为随着海拔增加先升高后降低的分布格局,依次为Ⅲ>Ⅱ>Ⅰ>Ⅳ,但蚤指数以Ⅳ垂直带最高(0.956 5),见表 4。
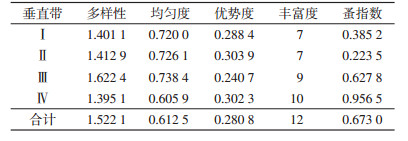
|
不同海拔垂直带小兽寄生蚤群落间相似性系数不同,其中以Ⅱ和Ⅲ垂直带最高(0.78),为极相似,其余垂直带两两之间均为中等相似,蚤类的组成存在一定差异,见表 5。
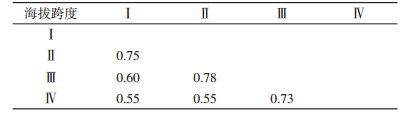
|
本次调查根据玉龙鼠疫疫源地植被分布特征,通过对小兽体表寄生蚤进行系统调查发现,玉龙鼠疫疫源地小兽体表寄生蚤优势种为方叶栉眼蚤、特新蚤指名亚种和棕形额蚤指名亚种。不同生境中蚤类的组成和分布有明显差异,耕地小兽寄生蚤数量多于林地,其中耕地的特新蚤指名亚种和棕形额蚤指名亚种的数量明显高于林地。
蚤的数量随着海拔梯度的增加呈现升高趋势,尤其在Ⅳ垂直带,蚤类总数或单个优势蚤种数量均远远超过前3个海拔梯度带蚤数量总和,反映该疫源地Ⅳ垂直带的整体环境有利于蚤类的生存与繁殖,寄生蚤始终维持在较高水平,可能有利于当地鼠疫的流行和暴发。特新蚤指名亚种在4个梯度均为优势种,从数量和生态幅水平均凸显其作为该疫源地重要传播媒介的地位。
宿主动物是成蚤的食物源,同时也是其重要生存环境因素,决定了蚤类对宿主动物的高度依赖性[12]。小兽体表寄生蚤的染蚤率和蚤指数较高是该疫源地一个主要特征。方叶栉眼蚤是大绒鼠的主要寄生蚤,特新蚤指名亚种和棕形额蚤指名亚种是齐氏姬鼠的优势种,体现了重要媒介对重要宿主动物的专寄性。
物种多样性指数是反映群落多样性高低的定量指标,在一定程度上反映了群落的复杂及稳定程度[13],玉龙鼠疫疫源地各垂直带总体呈现出蚤类物种多样性指数不高而均匀度指数较高的分布格局,说明该地区蚤类群落结构相对简单。另外,从蚤类物种多样性和均匀度随着海拔增加先升高后降低的变化趋势来看,媒介蚤类的空间分布均表现一定程度的聚集性分布,聚集分布格局意味着疫源地蚤类在不同海拔梯度和生境的地理分布不均匀,鼠疫易发生在Ⅳ垂直带农耕地或森林坡地,说明海拔和生境等环境因素决定疫源地鼠疫动物宿主和媒介生物的地理分布特征,并直接影响鼠疫的流行和传播模式。
近年来,根据鼠疫诊断标准证实动物鼠疫的流行情况显示,当地疫点主要在木梳、鹿子村、满下、满中、满上、旦都前、旦都后和汝南化3、4自然村社。对应相应的海拔梯度分布,除汝南化位于Ⅱ垂直带,其余疫点均分布于Ⅳ垂直带[14]。结合寄生蚤以Ⅳ垂直带的物种多样性和均匀度指数较低而优势度较高、总蚤指数最高等情况,认为该疫源地的Ⅳ垂直带是其核心区域,应作为鼠疫监测和防治的重点区域。
志谢: 本次调查在现场实施过程中得到玉龙县CDC的大力支持,特此志谢| [1] | 宋志忠, 夏连续, 梁云, 等. 云南玉龙及古城区鼠疫自然疫源地判定及初步研究[J]. 中国地方病防治杂志, 2008, 23(1): 3–7. |
| [2] | 李贵昌, 蔡文凤, 张福新, 等. 云南省玉龙鼠疫疫源地宿主及媒介监测结果分析[J]. 疾病监测, 2009, 24(2): 114–117. |
| [3] | 郭英, 韦蝶心, 梁云, 等. 云南省不同型鼠疫自然疫源地鼠疫菌生化特性比较[J]. 中华地方病学杂志, 2009, 28(4): 373–375. |
| [4] | 申小娜, 王琪, 夏连续, 等. 我国新发现的鼠疫自然疫源地鼠疫菌基因组序列测定及分析[J]. 中华地方病学杂志, 2011, 30(5): 476–480. |
| [5] | 朱俊洁, 王鹏, 李伟, 等. 云南省鼠疫菌株差异片段基因分型及其流行病学特征[J]. 中华地方病学杂志, 2013, 32(6): 599–601. |
| [6] | 张正飞, 张福新, 杨焕, 等. 丽江野鼠鼠疫自然疫源地指示动物血清学调查研究[J]. 中国地方病防治杂志, 2014, 29(1): 19–20. |
| [7] | 俞东征. 鼠疫动物流行病学[M]. 北京: 科学出版社, 2009: 112-139. |
| [8] | 解宝琦, 曾静凡. 云南蚤类志[M]. 昆明: 云南科技出版社, 2000: 110-315. |
| [9] | 詹银珠, 郭宪国, 左小华, 等. 云南省部分地区地里纤恙螨分布调查[J]. 中华流行病学杂志, 2011, 32(1): 13–16. |
| [10] | 中国科学院生物多样性委员会. 生物多样性研究的原理与方法[M]. 北京: 中国科学技术出版社, 1994: 141-165. |
| [11] | 徐正会, 杨比伦, 胡刚. 西双版纳片断山地雨林蚁科昆虫群落研究[J]. 动物学研究, 1999, 20(4): 288–293. |
| [12] | 龚正达, 吴厚永, 段兴德, 等. 云南横断山区蚤类物种多样性的地理分布趋势与重要环境因素的关系[J]. 生物多样性, 2001, 9(4): 319–328. |
| [13] | Schenier SM. Measuring pattern diversity[J]. Ecology, 1992, 73(5) : 1860–1867 .DOI:10.2307/1940037. |
| [14] | 张正飞, 张福新, 和映天, 等. 丽江野鼠鼠疫疫源地6年疫情发生监测及防治措施研究[J]. 国外医学医学地理分册, 2012, 33(3): 154–158. |