 2016, Vol. 27
2016, Vol. 27扩展功能
文章信息
- 姚丹丹, 隋晶晶, 冯志勇
- YAO Dan-dan, SUI Jing-jing, FENG Zhi-yong
- 湛江市褐家鼠种群年龄划分及繁殖特征
- The population age and reproductive characteristics of Rattus norvegicus in Zhanjiang city
- 中国媒介生物学及控制杂志, 2016, 27(5): 454-458
- Chin J Vector Biol & Control, 2016, 27(5): 454-458
- 10.11853/j.issn.1003.8280.2016.05.008
-
文章历史
- 收稿日期: 2016-04-22
- 网络出版时间: 2016-08-11




近年来,随着农村城镇化发展和新农村建设的推进,褐家鼠(Rattus norvegicus)种群数量大幅增加,为城镇和农舍鼠类的51.92%[1],威胁居民健康和农业生产。年龄的鉴定是研究动物种群生态学的重要基础[2],分析种群年龄结构有助于了解种群数量动态发展规律,对鼠害的预测预报有重要作用。国内外学者关于褐家鼠不同年龄的鉴定方法做了大量工作[3-4],体重法和胴体重法为最常用的划分指标[5]。由于鼠类个体大小受地理位置、生态环境和食物的影响较大,褐家鼠的年龄划分标准因地而异[6-7]。广东省湛江地区目前尚无褐家鼠种群生态特征的研究报道,该文以体重作为划分褐家鼠年龄组的标准,并探讨其与胴体重、体长和尾长的关系,分析各年龄组间繁殖力的差异。
1 材料与方法2008-2011年于湛江市的菜市场、垃圾场、房舍等褐家鼠密度高区域,定期采用笼捕法捕获褐家鼠,带回实验室备用。用乙醚麻醉后,称量体重、胴体重,测量体长、尾长等指标,解剖并观察其生殖器官发育和繁殖状况,计算种群睾丸下降率、睾丸大小和重量,雌鼠妊娠率、胎仔数及繁殖指数等。
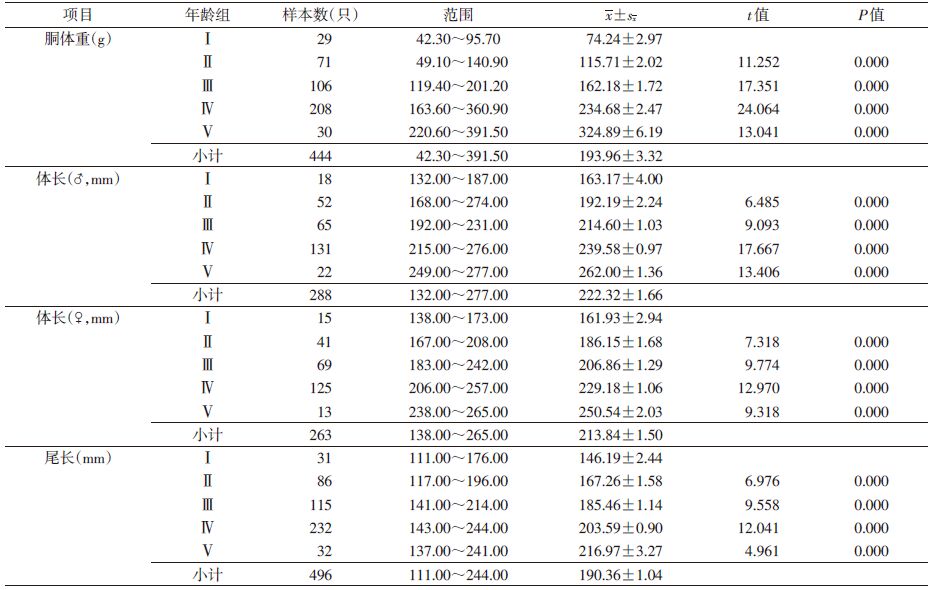
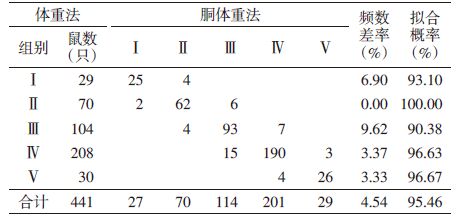
2 结果 2.1 年龄组的划分共捕获褐家鼠598只(♀275只,♂323只),对胴体重、体重、体长和尾长进行t检验,雌雄褐家鼠除体长外其他指标差异均无统计学意义,见表 1,故合并分析。
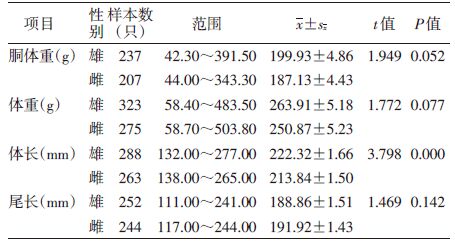
|
以每10.0 g体重为一个单位,将所有样本的体重进行频次分配,见图 1,根据褐家鼠的体重频次分布特征并参照繁殖状况,可将褐家鼠划分为5个年龄组:幼年组(Ⅰ,体重≤120 g),雌鼠无生殖活动迹象(妊娠率和繁殖指数均为0),雄鼠睾丸下降率低至15.79%;亚成年组(Ⅱ,体重120~180 g),开始进入性成熟期,4.88%雌鼠参与繁殖(均以肉眼可见有胚胎或子宫斑为准),雄鼠睾丸下降率为60.38%;成年Ⅰ组(Ⅲ,体重180~240 g),43.66%的雌鼠参与繁殖,雄鼠睾丸下降率为86.57%;成年Ⅱ组(Ⅳ,体重240~390 g),参与繁殖的雌鼠占90.15%,雄鼠睾丸下降率为94.81%;老年组(Ⅴ,体重>390 g),所有雌鼠均参与繁殖,雄鼠睾丸下降率为100%。相邻两年龄组间体重经t检验,差异有统计学意义(P=0.000),见表 2。体重随着种群年龄的增长而增加,说明该标准可用于鉴定褐家鼠年龄。

|
| 图 1 湛江市褐家鼠的体重频次分布 Figure 1 The weight frequency distribution of R. norvegicus |
| |
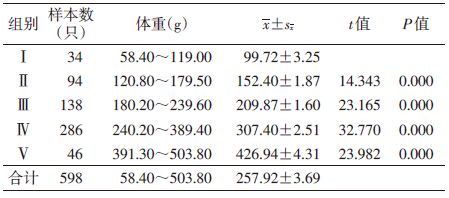
褐家鼠尾长短于体长,为体长的87.21%。经t检验,不同年龄组相邻两组间胴体重、体长与尾长间差异均有统计学意义(P=0.000),且随着种群年龄的增长,胴体重、体长和尾长不断增加,见表 3。
|
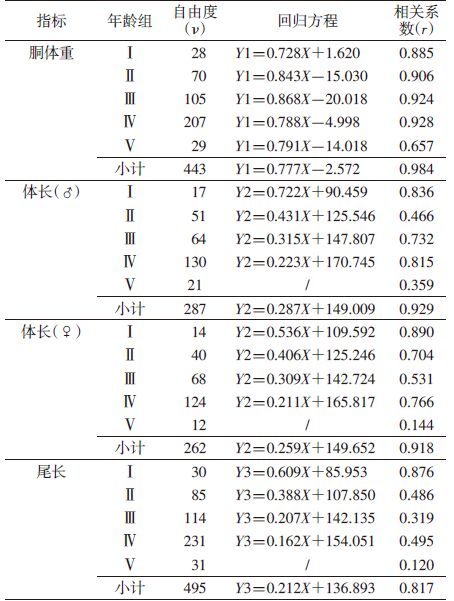
5个年龄组总体重(X)与胴体重(Y1)呈正相关(r=0.984);各年龄组显著相关。各年龄组回归方程:Ⅰ. Y1=0.728X+1.620,Ⅱ. Y1=0.843X-15.030,Ⅲ. Y1=0.868X-20.018,Ⅳ. Y1=0.788X-4.998,Ⅴ. Y1=0.791X-14.018(表 4)。通过回归方程推算,5个年龄组相应的胴体重标准为幼年组胴体重<89.0 g,亚成年组89.0~136.0 g,成年Ⅰ组136.1~188.0 g,成年Ⅱ组188.1~302.0 g,老年组>302 g。
|
对441只褐家鼠按体重法及胴体重法划分的年龄组进行检验分析,其频数分布大部分重合,5组鼠数相差值分别为2、0、10、7和1,按鼠总数(n=441)计算频数差率(│A-B│/A×100%,A为按体重法、B为按胴体重法划分的某一年龄组的鼠数),总计仅为4.54%,拟合概率达95.46%;即使按原体重法划分的各组鼠数计算频数差率,各年龄组均<10%,个别组频数差为0,见表 5。两种方法测定的各年龄组在种群中所占数量均比较接近,总体拟合度较高,故实际工作中2种方法均可采用。
|
雌雄褐家鼠的体长有差异,故分开计算。雌雄总体长与体重显著相关〔Y2=0.287X+149.009(♂)、Y2=0.259X+149.652(♀)〕(表 4)。但仅前4组的体长与体重显著相关,说明前4组体长随体重的增加而增长,老年后体长几乎停止增长,故不宜采用体长划分褐家鼠年龄组。
2.3.3 体重与尾长的相关性总体尾长与体重显著相关(Y3=0.212X+136.893)(表 4),但与体长一样,仅前4组显著相关。说明从幼年组到成年Ⅱ组,尾长与体重同步增长,到老年后,尾长的增长滞后于体重的增加,因此在划分年龄组时,尾长仅可作为参考性指标,见表 4。
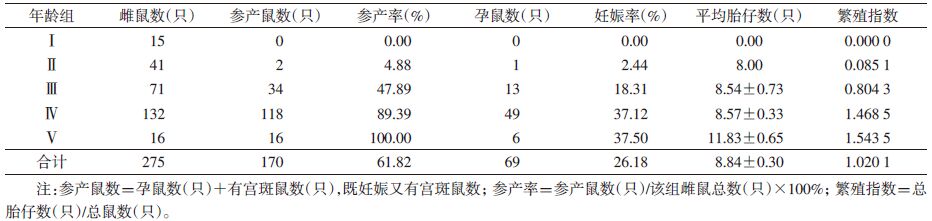
2.4 不同年龄组种群繁殖特征 2.4.1 雌鼠繁殖特征275只褐家鼠雌鼠繁殖参数,雌鼠幼年组均为无孕个体;亚成年组有2只参与繁殖,参产率为4.88%,孕鼠1只,妊娠率为2.44%,孕鼠体重为155.3 g,平均胎仔数为8只;成年Ⅰ组妊娠率迅速增加,至老年组时最高,说明其成年组和老年组繁殖能力最强。参产率由幼年组的0逐渐增加到老年组的100%,说明老年组雌鼠均参与繁殖。在老年组的6只孕鼠中,胎仔数为9~14只,平均胎仔数(11.83±0.65)只,远高于其他年龄组的平均胎仔数,与成年组比较差异有统计学意义(t=3.301,P=0.002),见表 6,而其他组胎仔数最多13只。随种群年龄的增长繁殖指数升高,雌鼠繁殖力不断增强,褐家鼠种群繁殖的主体为成年组和老年组。
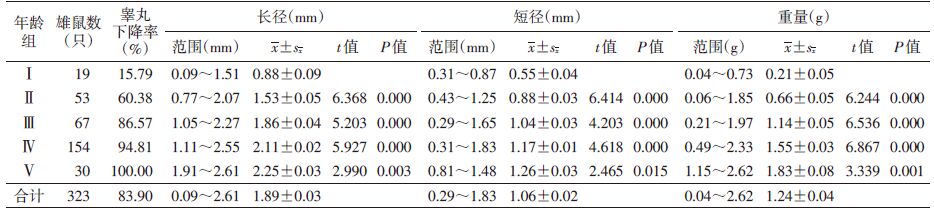
随着年龄增加睾丸下降率逐渐升高,幼年组有少量个体性成熟,睾丸开始下降,下降率为15.79%,雄鼠最小体重为69.5 g;亚成年组部分个体性成熟,睾丸下降率为60.38%;成年Ⅰ组、成年Ⅱ组和老年组睾丸下降率达86.57%~100%,为亚成年组的1.43~1.66倍,平均睾丸下降率为93.23%,不同年龄组雄鼠睾丸下降率差异有统计学意义(χ2=100.228,P<0.01)。雄鼠睾丸的长径、短径和重量随年龄增长而增加,相邻两组间经t检验,差异均有统计学意义(P=0.000),表明褐家鼠的睾丸随年龄增长逐渐增大,见表 7。
褐家鼠体重随着年龄的增长而增加,各组间差异均有统计学意义,胴体重和体重在全部样本及各年龄组中均呈显著正相关,因此两者均可作为年龄划分的重要指标。与臼齿磨损度[8-9]、晶体干重[9]、头骨形态[10]等指标相比,体重法简便易行,更适用于基层操作[11-13]。而胴体重法可排除繁殖及摄食状况对年龄鉴定的影响,优于体重法[14]。
体重法存在较大的地区差异,姚伟兰等[15]比较了广州和深圳市2个饲养场褐家鼠种群的生态特征差异,尽管两种群的地理位置和食物资源差异较小,但是相同年龄组的体重范围差异较大,两种群除幼年组外其他各年龄组平均体重均大于本研究相同年龄组的体重,因此研究某一地区鼠类生态特征首先应制定该地区鼠类年龄的划分标准。
3.2 褐家鼠的繁殖特征啮齿动物的繁殖特征是动物与环境长期互相作用的结果。同一地区的同一鼠种在不同环境条件下,其胎仔数和繁殖强度存在差别。本研究中褐家鼠每胎平均胎仔数为8.84只,与张涛等[16]报道结果(5.35只)存在较大差异,可能与栖息环境和食物来源有关。
褐家鼠不同年龄组间种群繁殖力存在显著差异,繁殖力随着年龄的增长而增强,成年组和老年组为种群繁殖的主体。老年组平均胎仔数远高于其他年龄组的平均胎仔数,老年组胎仔数多、妊娠率高的现象在其他鼠种中亦有报道[17-19],目前主要有两种解释,一是为补偿老年组总体繁殖力下降的一种生理适应性[17],二是害鼠进入老年组后死亡率很高,而剩余真正的衰老个体较少,因而仍保持着较强的繁殖力。
| [1] | 姚丹丹, 隋晶晶, 刘明津, 等. 农村城镇化对鼠类群落结构的影响研究[J]. 中国媒介生物学及控制杂志,2014,25 (6) :518–520. |
| [2] | 陈立军, 刘伟, 苏永志, 等. 典型草原区达乌尔鼠兔年龄划分标准[J]. 动物学杂志,2013,48 (3) :345–350. |
| [3] | Yabe T. Eye lens weight as an age indicator in Norway rat[J]. J Mammal Soc Jap, 1979, 8 (1) : 54–55 . |
| [4] | 陈荣海, 董志刚, 杨春文. 应用阴茎骨鉴定雄性褐家鼠年龄组的探讨[J]. 兽类学报,1988,8 (4) :288–293. |
| [5] | 李世斌, 陈安国, 李波, 等. 洞庭平原褐家鼠年龄分组及种群年龄动态分析[J]. 兽类学报,1993,13 (2) :123–130. |
| [6] | 颜赛勋, 张艺, 马合木提·哈力克, 等. 新疆褐家鼠种群生命表及其在干旱区的生存能力分析[J]. 生物学杂志,2006,23 (1) :31–33. |
| [7] | 洪朝长. 褐家鼠种群年龄组成和种群动态预测[J]. 中国鼠类防制杂志,1987,3 (3) :133–136. |
| [8] | 邹波, 王宇翔, 郭永旺, 等. 山西省娄烦县北社鼠种群年龄结构及季节变动研究[J]. 中国媒介生物学及控制杂志,2013,24 (1) :31–33. |
| [9] | 郭志南, 苏成豪, 陈国伟, 等. 黄毛鼠种群年龄划分的研究[J]. 中华卫生杀虫药械,2014,20 (4) :332–335. |
| [10] | 张碧岱, 邹波, 花慧贞. 山西省娄烦县中华鼢鼠种群年龄结构研究[J]. 山西科技,2013,28 (5) :38–40. |
| [11] | 杨再学, 雷邦海, 潘世昌, 等. 贵州省大足鼠种群年龄鉴定和繁殖参数变化[J]. 山地农业生物学报,2015,34 (4) :39–42. |
| [12] | 张世炎, 陈安. 板齿鼠种群年龄的研究[J]. 中华卫生杀虫药械,2013,19 (2) :138–140. |
| [13] | 张世炎, 陈安. 海南屋顶鼠种群年龄的研究[J]. 医学动物防制,2012,28 (3) :237–238. |
| [14] | 李秋阳, 赵秀兰, 杨滨. 云南省沧源县黄胸鼠种群年龄组的划分及分析[J]. 中国媒介生物学及控制杂志,2013,24 (1) :39–42. |
| [15] | 姚伟兰, 戚根贤, 王骏, 等. 褐家鼠种群生态特征的研究[J]. 中山大学学报论丛,1998, (4) :89–93. |
| [16] | 张涛, 冯志勇, 陈戊申, 等. 广东省乡镇家栖鼠形动物种类结构及季节繁殖的观察[J]. 中国媒介生物学及控制杂志,2009,20 (4) :295–297. |
| [17] | 张美文, 黄璜, 王勇, 等. 洞庭湖区社鼠的繁殖生态[J]. 生态学报,2006,26 (3) :884–894. |
| [18] | 杨再学, 金星, 郭永旺, 等. 应用胴体重鉴定高山姬鼠种群年龄[J]. 山地农业生物学报,2011,30 (2) :104–109. |
| [19] | 杨再学, 雷邦海, 郑元利, 等. 针毛鼠种群年龄组的划分标准及其繁殖力[J]. 贵州农业科学,2014,42 (10) :164–166. |