 2016, Vol. 27
2016, Vol. 27扩展功能
文章信息
- 鲁亮, 蒋卫, 刘蓉蓉, 黎唯, 朱志勇, 帕尔哈提, 波拉提, 王钦琰, 陈志刚, 刘云翔, 刘京利, 安文严, 冯玉明, 甫拉提, 帕尔哈提, 杨庭祥, 胡俊, 鲜军, 刘起勇
- LU Liang, JIANG Wei, LIU Rong-rong, LI Wei, ZHU Zhi-yong, Paerhati, Bolati, WANG Qin-yan, CHEN Zhi-gang, LIU Yun-xiang, LIU Jing-li, AN Wen-yan, FENG Yu-ming, Fulati, Paerhati, YANG Ting-xiang, HU Jun, XIAN Jun, LIU Qi-yong
- 中国-哈萨克斯坦边境地区啮齿动物多样性调查
- Investigation on rodent diversity along the China-Kazakhstan border area
- 中国媒介生物学及控制杂志, 2016, 27(4): 317-320
- Chin J Vector Biol & Control, 2016, 27(4): 317-320
- 10.11853/j.issn.1003.8280.2016.04.001
-
文章历史
- 收稿日期: 2016-05-08
- 网络出版时间: 2016-06-03 12:10



2 新疆维吾尔自治区疾病预防控制中心, 乌鲁木齐 830001;
3 博尔塔拉蒙古自治州疾病预防控制中心, 新疆 博乐 833400;
4 额敏县疾病预防控制中心, 新疆 额敏 834600;
5 精河县疾病预防控制中心, 新疆 精河 833300;
6 霍城县疾病预防控制中心, 新疆 霍城 835200;
7 温泉县疾病预防控制中心, 新疆 温泉 833500;
8 克拉玛依市疾病预防控制中心, 新疆 克拉玛依 834000
2 Xinjiang Center for Disease Control and Prevention;
3 Boertala Center for Disease Control and Prevention;
4 Emin Center for Disease Control and Prevention;
5 Jinghe Center for Disease Control and Prevention;
6 Huocheng Center for Disease Control and Prevention;
7 Wenquan Center for Disease Control and Prevention;
8 Kelamayi Center for Disease Control and Prevention
哈萨克斯坦是我国在中亚地区的重要邻国,两国边境线超过1 500 km,从天山山脉绵延到阿尔泰山脉。中国-哈萨克斯坦边境(中哈边境)地区,历史上是“丝绸之路”的重要通道,现在是中国和中亚地区物流和能源的重要通道。目前,中哈两国已无领土争端,边境地区相对稳定,在“一带一路”的建设和开发中,该边境地区将成为新疆维吾尔自治区经济建设的重要区域。中哈边境地区地形复杂,生境多样,多为雪山、高山森林、荒漠、草原和湿地,为啮齿动物提供了各种栖息地。因此,该地区以啮齿动物为代表的小兽种类十分丰富[1],存在多种类型鼠疫自然疫源地。天山山脉有灰旱獭(Marmota baibacina)和长尾黄鼠(Spermophilus parryii)鼠疫疫源地;准噶尔盆地有大沙鼠(Rhombomys opimus)鼠疫自然疫源地[2],威胁当地经济发展。
夏尔西里地区位于博尔塔拉州,海拔1 210~3 670 m。高海拔地区以高山针叶林为主,中低海拔则以温带草原、阔叶林和灌丛为主[3-4]。由于此地之前属于军事争议区,因此自然生境保持完好,划属我国后为国家级自然保护区。曾有关于植被、重要病媒感染病原体的初步调查,但比较简单[3, 5],至今未系统调查区域内的鼠类。由于博尔塔拉州境内分布有两种鼠疫疫源地,通过了解夏尔西里地区鼠种的构成,可以判断当地可能受到的鼠疫威胁,以便开展健康教育工作。准确鉴定和分析当地啮齿动物种类和群落构成,有利于动物间鼠疫疫情的动态研究及掌握流行机制。本研究在中哈边境多个地点采集了以啮齿动物为主的小型兽类,通过DNA条形码技术准确鉴定种类[6],为了解当地啮齿动物群落构成提供重要数据。
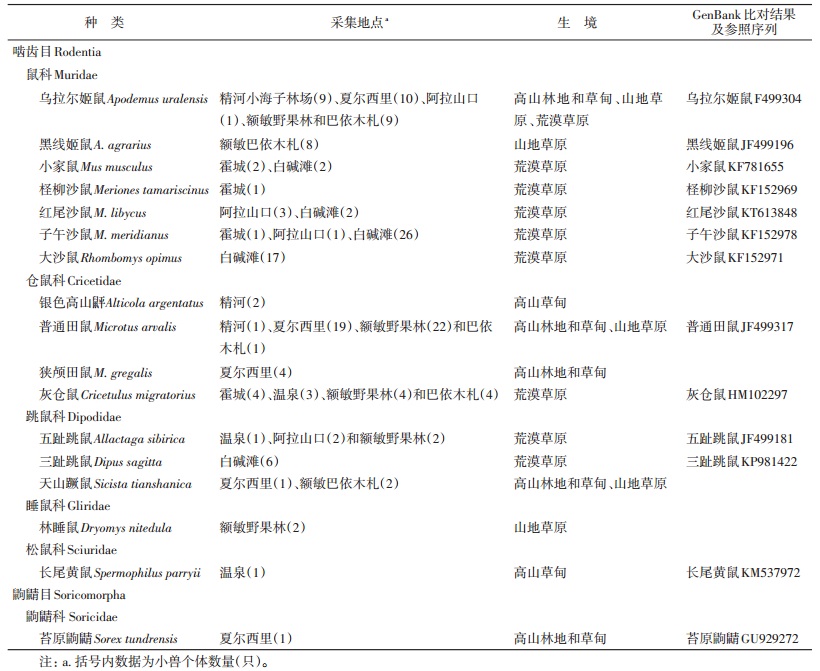
1 材料与方法 1.1 样品采集于精河县小海子林场、霍城县、温泉县、夏尔西里、阿拉山口、额敏县野果林和巴依木札、克拉玛依白碱滩采集标本,小兽种类、数量及生境分布见表 1。
|
在现场首先通过小兽外部形态特征进行鉴定[1],根据其外部形态无法确定的标本则制作剥制标本,观察外部和头骨形态,并测量相关数据,通过检索表鉴定。
1.3 DNA条形码扩增和分析 1.3.1 模板DNA提取使用Qiagen DNA提取试剂盒提取肝脏或肌肉DNA,4℃保存备用。
1.3.2 PCR扩增与测序扩增时首先选用通用引物(BatL5310,R6036R)[7]扩增细胞色素C氧化酶亚基Ⅰ(COⅠ)基因片段。当通用引物扩增失败时,则选用鸡尾酒引物(VF1LFt1、VR1LRt1)[8]。10×缓冲液2.5μl,dNTP 0.5μl,引物各0.5μl,LA Taq DNA聚合酶0.15μl,模板DNA 1μl,ddH2O 19.85μl。PCR反应条件:95℃预变性5 min,95℃变性45 s,54℃退火1 min,72℃延伸1 min,35个循环,72℃延伸10 min。扩增产物经1%琼脂糖凝胶电泳检测,送测序公司(北京天一辉远生物科技有限公司)进行双向测序。
1.3.3 DNA序列分析测序所得的序列文件经过Mega 6.0软件中的MUSCLE模块进行比对排列,然后使用Neighbor?Joining方法构建系统树,分析个体间的遗传距离和关系。系统树中每个末端分支代表样本的基因在GenBank中进行Blast比对,选择形态鉴定符合或相似,比对结果相似度高于98%的序列作为参照序列。
2 结果 2.1 形态鉴定结果在中哈边境的7个市(县)共采集鼠类和鼩鼱174只。形态鉴定共17种,其中鼠科7种,仓鼠科4种,跳鼠科3种,睡鼠科1种,松鼠科1种,鼩鼱目的鼩鼱科1种。
2.2 分子鉴定结果通过DNA条形码测序和系统树构建,进一步核对形态鉴定结果。DNA条形码的Neighbor-Joining系统树分为17个高度支持的分支。说明根据形态特征的分类基本准确,但普通田鼠和狭颅田鼠的数量发生变化。由于田鼠属的外部形态非常相似,根据外部形态很难区分。经过DNA条形码的比对,发现47份田鼠标本主要以普通田鼠为主,狭颅田鼠仅有4份。
通过与GenBank中序列的Blast比对,发现12个种的序列可在GenBank中找到一致的序列,进一步证实了形态学的鉴定结果。形态鉴定为子午沙鼠的标本,其DNA条形码序列与GenBank中的郑氏沙鼠(Meriones zhengi)序列具有更高的相似度,与我国内蒙古、河北省等地的子午沙鼠存在较大的遗传距离。狭颅田鼠、银色高山鼠平、天山蹶鼠和林睡鼠在GenBank中均无相似度>96%的序列,形态学分类结果见图 1。

|
| 图 1 基于Kimrua 2-parameter模型构建的中哈小型兽类Neighbor-Joining系统关系树 Figure 1 A Neighbor?Joining tree of small mammals collected from China-Kazakhstan border area |
| |
本次调查获得较多的种类有普通田鼠、灰仓鼠、大沙鼠、郑氏沙鼠、乌拉尔姬鼠和五趾跳鼠。通过分析种内个体的遗传距离,多为0~1.1%,远小于种间个体遗传距离。
3 讨论本次调查覆盖的区域以3种生境为主,即山地森林草甸草原(精河县小海子林场、夏尔西里和额敏县野果林)、荒漠草原(霍城、温泉、阿拉山口和克拉玛依的白碱滩)和山地草原(额敏县巴依木札)。3种生境的啮齿动物构成完全不同。在山地森林草甸草原,采集到乌拉尔姬鼠、普通田鼠、狭颅田鼠、银色山鼠平、天山蹶鼠、林睡鼠、长尾黄鼠和苔原鼩鼱;在荒漠草原,采集到小家鼠、灰仓鼠、三趾跳鼠、五趾跳鼠、红尾沙鼠、柽柳沙鼠、郑氏沙鼠、大沙鼠和乌拉尔姬鼠;山地草原中则采集到乌拉尔姬鼠、黑线姬鼠、普通田鼠、灰仓鼠和天山蹶鼠。
夏尔西里地区为军事控制区,人类活动和干扰较少,保持良好的自然状态。林地主要分布在阴坡,高海拔地区以云杉为主,中低海拔地区以桦树为主。阳坡以草甸为主,多为一些水源相对丰富的山谷,灌木和杂草密集。在云杉林中,捕获到狭颅田鼠和苔原鼩鼱,但数量较少。在桦树林中以普通田鼠和乌拉尔姬鼠为主。山谷较高的杂草丛中捕获到天山蹶鼠,但数量很少。由于调查时间较晚且短,仅在夏尔西里的局部观察到黄鼠活动,但未捕获。根据夏尔西里的生境特点,黄鼠可能分布在高山草甸。
子午沙鼠是我国北方荒漠草原中常见的沙鼠,广泛分布于不同类型的沙鼠鼠疫疫源地。子午沙鼠在不同疫源地鼠疫菌的传播和保存过程中作用不同。在长爪沙鼠鼠疫疫源地,子午沙鼠是动物间鼠疫流行的积极参与、传播和保存者;而在大沙鼠鼠疫疫源地中,子午沙鼠却无相似的地位[2]。有研究发现我国的子午沙鼠存在2个基因型[9-10]。其中伊犁河谷和准噶尔盆地的子午沙鼠与我国其他地区的子午沙鼠遗传差异较大。
此次调查在霍城、阿拉山口和克拉玛依的白碱滩均捕获到形态接近子午沙鼠的标本。通过DNA条形码序列与GenBank中的数据进行比对,发现与采自新疆霍尔果斯的“郑氏沙鼠”有较高的相似度,而与其他地区的子午沙鼠有较大的遗传距离。鉴于GenBank中的“郑氏沙鼠”无明确的形态学依据[11],我们不认为本次采集的是“郑氏沙鼠”。但该标本与内蒙古自治区、青海地区的子午沙鼠差异较大,遗传距离>11%。根据DNA条形码数据在鼠类中的一般规律,COⅠ基因的遗传距离较大,应属于2个不同种。因此,本次调查认为伊犁河谷和准噶尔盆地的子午沙鼠为一个独立存在的沙鼠种。但与子午沙鼠的外部形态非常接近,该鼠种在新疆的分布及在大沙鼠鼠疫疫源地宿主动物群落中的地位有待进一步研究。
| [1] | 王思博, 杨赣源. 新疆啮齿动物志[M]. 乌鲁木齐: 新疆人民出版社, 1983 : 87 -210. |
| [2] | 俞东征. 鼠疫动物流行病学[M]. 北京: 科学出版社, 2009 : 237 -306. |
| [3] | 赵阳, 陈蜀江, 马静, 等. 夏尔西里自然保护区野生近缘植物研究[J]. 科技创新导报,2013, (13) :142–143. |
| [4] | 陈蜀江. 新疆夏尔希里自然保护区综合科学考察[M]. 乌鲁木齐: 新疆科学技术出版社, 2006 : 35 -80. |
| [5] | 刘晓明, 张桂林, 刘然, 等. 新疆维吾尔自治区夏尔西里自然保护区全沟硬蜱复合感染蜱媒病原研究[J]. 中华流行病学杂志,2015,36 (10) :1153–1157. |
| [6] | Hebert PDN, Gregory TR. The promise of DNA barcoding for taxonomy[J]. Syst Biol, 2005, 54 (5) : 852–859 .DOI:10.1080/10635150500354886. |
| [7] | Ivanova NV, Clare EL, Borisenko AV. DNA barcoding in mammals[M]/Kress WJ, Erickson DL. DNA Barcodes. Totowa NJ:Humana Press, 2012, 858:153-182. |
| [8] | Ivanova NV, Zemlak TS, Hanner RH, et al. Universal primer cocktails for fish DNA barcoding[J]. Mol Ecol Notes, 2007, 7 (4) : 544–548 . |
| [9] | Wang Y, Zhao LM, Fang FJ, et al. Intraspecific molecular phylogeny and phylogeography of the Meriones meridianus (Rodentia:Cricetidae) complex in northern China reflect the processes of desertification and the Tianshan Mountains uplift[J]. Biol J Linn Soc, 2013, 110 (2) : 362–383 .DOI:10.1111/bij.2013.110.issue-2. |
| [10] | Yazdi FT, Adriaens D, Darvish J. Geographic pattern of cranial differentiation in the Asian Midday Jird Meriones meridianus (Rodentia: Muridae: Gerbillinae)and its taxonomic implications[J]. J Zool Syst Evol Res, 2012, 50 (2) : 157–164 . |
| [11] | Yin YQ, Jiang W, Zhang Z, et al. The divergence of small mammals in Xinjiang, China, as revealed by phylogentic analyses of COⅠand Cytb[J]. Anim Biol, 2014, 64 (2) : 163–176 . |