 2016, Vol. 27
2016, Vol. 27扩展功能
文章信息
- 岳仁苹, 龚正达, 王海波, 葛军旗, 张丽云, 周红宁
- YUE Ren-ping, GONG Zheng-da, WANG Hai-bo, GE Jun-qi, ZHANG Li-yun, ZHOU Hong-ning
- 云南省澜沧江流域居民区蚊类γ-多样性空间梯度格局与环境因素关系
- Spatial distribution pattern of mosquitoes γ-diversity and relationship with environmental factors of residential area along Lancang river in Yunnan province
- 中国媒介生物学及控制杂志, 2016, 27(3): 220-227
- Chin J Vector Biol & Control, 2016, 27(3): 220-227
- 10.11853/j.issn.1003.8280.2016.03.003
-
文章历史
- 收稿日期: 2016-02-24
- 网络出版时间: 2016-04-14 12:00




2 云南省地方病防治所, 云南 大理 671000;
3 北京市朝阳区疾病预防控制中心, 北京 100121;
4 云南省寄生虫病防治所, 云南 普洱 665000
2 Yunnan Institute of Endemic Diseases Control and Prevention;
3 Chaoyang District Center for Disease Control and Prevention;
4 Yunnan Institute of Parasitic Diseases Control and Prevention
空间梯度变化是物种多样性大空间尺度格局的显著特征,包含沿纬度、海拔等梯度的格局与变化趋势。物种多样性大空间尺度格局与环境因子密切相关,如物种丰富度与年降雨量和年平均温度等。不同自然区域中各物理因子作用不同,中国的青藏高原地区以海拔为主,在干旱和半干旱地区以湿度为主[1-2]。然而,物种多样性空间梯度格局的解释仍存在较大争议[3-6],无具体环境因子或假说得到一致认同,多认为因不同研究、地理区域和空间尺度引起[5-10]。
云南省属亚热带湿润季风气候类型,地势南低北高。近几年,对滇西北“三江并流”区域的澜沧江、怒江和金沙江三江上游居民区蚊类物种多样性空间分布格局与地理因素关系进行研究,发现各流域因所处地理位置、雨量分布特征、气候环境条件和居民区周边农耕地类型不同,蚊类物种多样性的分布格局和特征均存在一定差异,并与环境因子和人为干扰相关[9-11]。研究认为,在同一山地的不同森林植被带的自然环境和条件下,或同一纬度带的不同山地或河谷间,蚊类区系和物种多样性分布是否反映类似的空间格局和变化规律,或其他的气候类型、地域或江河流域的多样性与区系分布的规律和特征如何,局域或个案研究的结果和特征是否具有代表性等问题,在更大空间尺度、气候及环境类型的条件下有待进一步调查研究[10]。为进一步了解我国西南部湿润地区蚊类物种多样性大尺度空间梯度格局、变化趋势、特征及与重要环境因素关系,在云南省澜沧江上游调查基础上[9],对云南省澜沧江全流域(21°~30°N、海拔500~3 500 m,全长约1 250 km)的居民区蚊类进行调查取样,并对不同纬度和海拔梯度带蚊类的物种γ-多样性(物种丰富度)、空间分布格局、地理分布趋势、区系分异及与重要地理环境因素关系等进行研究和探讨。
1 材料与方法 1.1 区域概况澜沧江源于中国青藏高原,流经中国、缅甸、老挝、泰国、柬埔寨和越南,系国际河流,在东南亚称为湄公河,是世界第六大河,亚洲第二大河。澜沧江-湄公河全长4 880 km,在我国内河道里程2 161 km。贯穿云南省西部,跨越9个纬度梯度带(21°~30°N),共1 247 km,流经区域因地理位置不同和高度差,从南向北或由低到高依次出现低山、中山、亚高山、高山和极高山,或热带、亚热带、温带和寒带不同的气候带和森林植被景观,全流域地处热带和亚热带高原季风气候区。云南省澜沧江流域居民区的分布范围最高处位于德钦县升平(海拔约3 500 m),最低为勐腊县澜沧江与南腊河交汇处(海拔480 m)。与我国其他地区不同,该地区蚊类多样性较高和物种分布广[12-17]。
1.2 调查取样方法γ-多样性指区域或大陆尺度的物种数量,其分布格局与地理演化历史、地理隔离及环境梯度有较大关系。研究物种沿环境梯度的变化格局时,一般以一定的环境区间内出现的物种数为研究对象,包含γ-多样性研究的内涵。因此,γ-多样性沿环境梯度格局的变化主要体现在相同地理条件下沿环境梯度的变化规律和特征。
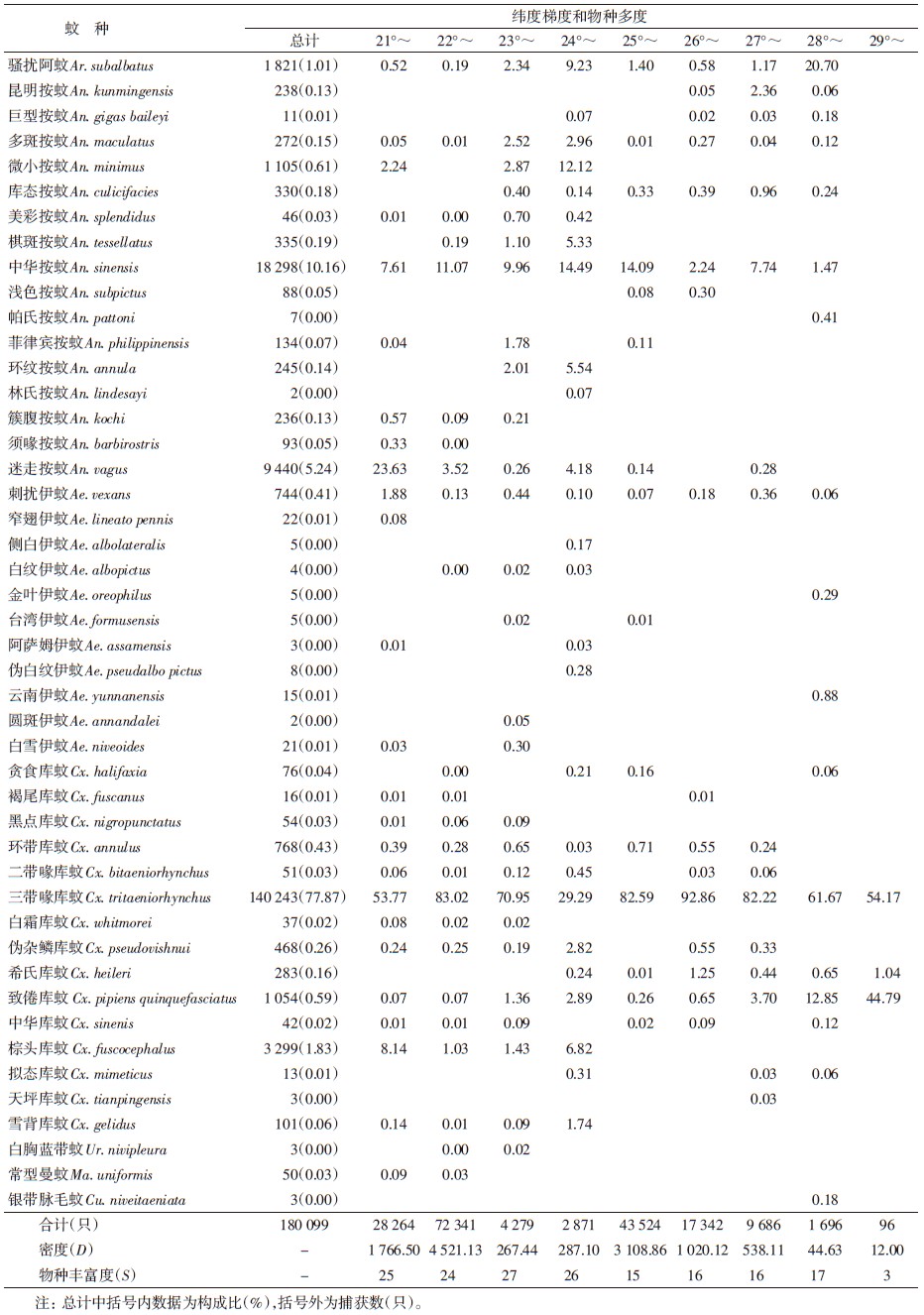
2005和2007年于云南省雨季、蚊类密度的高峰期7-9月进行调查。将云南省境内澜沧江流域(北纬21°~30° N、海拔500~3 500 m)作为调查的空间范围。空间梯度的划分:按每1° (60′)作为水平梯度带,从南到北调查可划分为9个带,以Ⅰ~Ⅸ表示,21°~(Ⅰ)、22°~(Ⅱ)、23°~(Ⅲ)、24°~(Ⅳ)、25°~(Ⅴ)、26°~(Ⅵ)、27°~(Ⅶ)、28°~(Ⅷ)和29°~(Ⅸ);调查区按每500 m划分垂直梯度带,分6个带,以A~F依次表示。以乡村居民区夜间活动的成蚊为研究对象,把每半个纬度(30′,直线约55 km)的距离为取样区间,在每个取样区间内选择不同海拔高度的2~3个乡镇及多个村庄作为调查点,采用灯诱捕法取样,应用UV诱蚊灯(12 V,300 mA,武汉市吉星环保科技有限责任公司生产),每个调查点置灯4~5盏,对居民点庭院、畜房夜间活动的成蚊进行通宵取样(19:30-07:30),每个点各捕蚊1夜。所获标本带回当地疾病预防控制中心实验室冰冻至死后,分类鉴定、计数和记录。同期收集当地纬度、海拔、平均气温和降雨量等相关信息。
1.3 数据处理及分析以各纬度和海拔梯度内出现的物种数表示物种丰富度,描述和比较γ-多样性(物种丰富度)沿纬度、海拔梯度带的变化规律;用β-多样性(Cody指数)比较相邻纬度或海拔梯度带间物种组成差异;用组平均系统聚类法[18]分析不同梯度带物种的区系分异及其关系。
(1)γ-多样性:

|
(2)β-多样性指数:
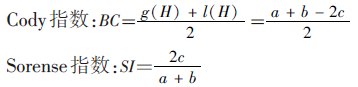
|
(3)蚊类密度(D)=样区捕蚊总数/(诱蚊灯数×夜晚数)
(4)物种多度(Abundance)(各蚊种数量的构成比):
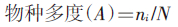
|
公式中,S为各梯度带的物种数,a和b分别为两梯度带各自的物种数,c为两梯度带的共有物种数,g(H)为沿梯度带增加的物种数,l(H)为沿梯度带减少的物种数,ni为每一物种个体数,N为总个体数。
(5)相关分析:应用SPSS 13.0软件对澜沧江流域所有取样点蚊类物种丰富度与密度、纬度、海拔、温度和降雨量做多元相关分析。
2 结 果 2.1 蚊类的组成及分布通过对云南省西部澜沧江流域12个县(市)30个乡(镇)居民区的调查,共捕获蚊类180 099只,隶属2亚科7属46种。不同纬度和海拔梯度带调查结果见表 1、2。其中按蚊属(Anopheles)16种,库蚊属(Culex)15种,伊蚊属(Aedes)11种,蓝带蚊属(Uranotaenia)、阿蚊属(Armigeres)、曼蚊属(Mansonia)和脉毛蚊属(Culiseta)各1种,见表 1。所获蚊类中,三带喙库蚊(Cx. tritaeniorhynchus)和致倦库蚊(Cx. pipiens quinquefasciatus)在各个取样点均有分布,分布最广;中华按蚊(An. sinensis)、多斑按蚊(An. maculatus)、刺扰伊蚊(Ae. vexans)、环带库蚊(Cx. annulus)及骚扰阿蚊(Ar. subalbatus)在大部分取样点有分布,分布较为广泛;其他蚊种分布范围相对较窄。三带喙库蚊占捕获蚊类总数的77.87%,其物种相对多度在各纬度梯度带和海拔梯度带占绝对优势,为29.29~92.86和64.29~86.83,是澜沧江流域居民区蚊类的优势种;中华按蚊占10.16%;迷走按蚊(An. vagus)、棕头库蚊(Cx. fuscocephalus)和骚扰阿蚊分别占5.24%、1.83%和1.01%,是该区域的常见种;其他42种蚊类占捕获总数均<1%,为该区域少见种类。
 |
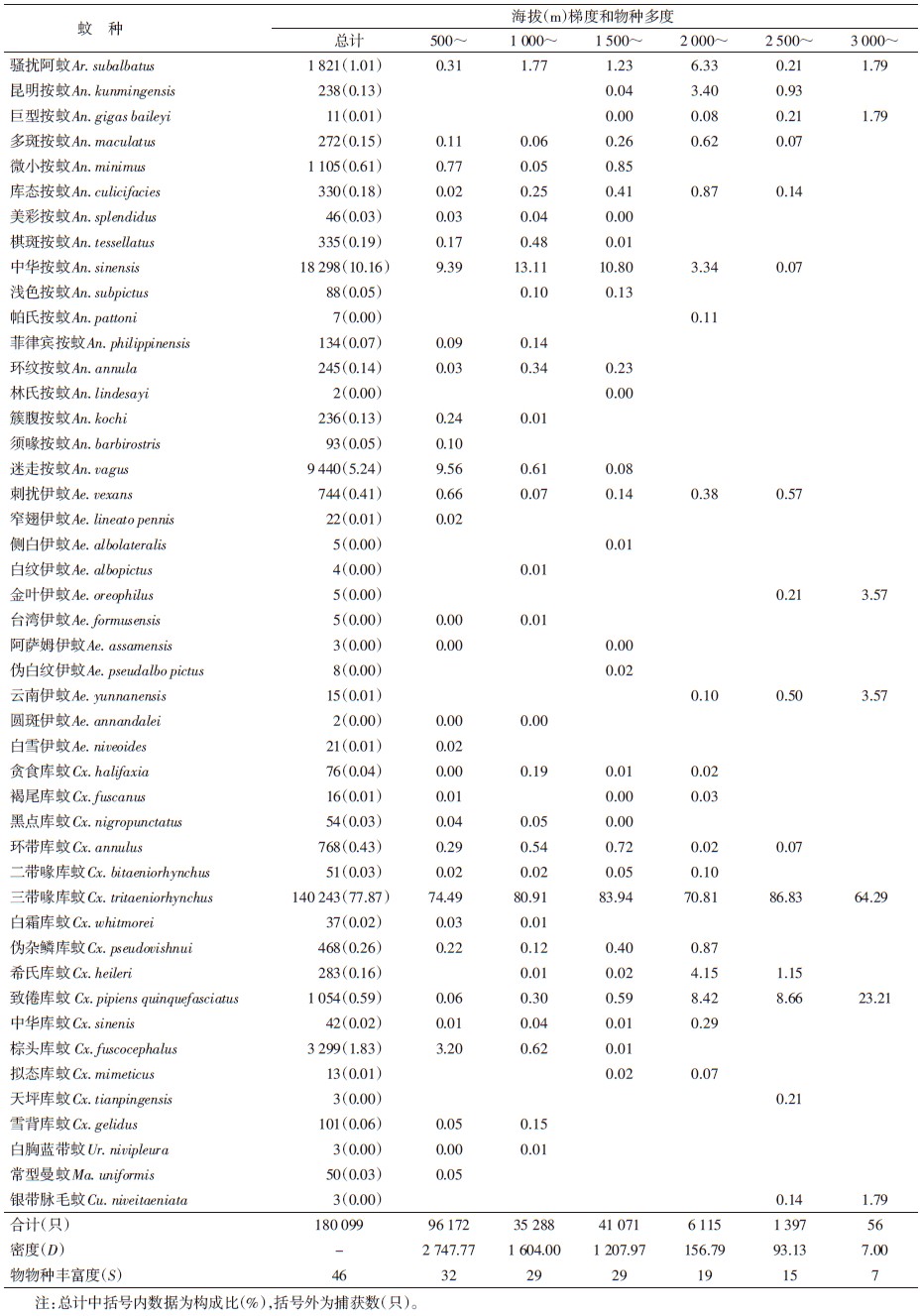 |
蚊类物种丰富度在低纬度21°~24°时较高,达24~27种,各梯度间物种丰富度变化较小(表 1、图 1)。但物种丰富度随纬度的升高减少,29°时最少,仅3种蚊类。蚊类物种丰富度在低海拔区域较高,峰值在500~1 000 m,达32种。但随着海拔的升高,物种丰富度整体也呈现逐渐下降的趋势(表 2、3)。在中海拔>2 000 m区域,蚊类丰富度下降明显,至高海拔3 000~3 500 m区间蚊种组成较为简单,仅发现7种。综上所述,澜沧江流域居民区蚊类γ-多样性的空间分布格局总体呈现随纬度和海拔的增高而递减趋势。

|
| 注:Ⅰ(21°~); Ⅱ(22°~); Ⅲ(23°~); Ⅳ(24°~); Ⅴ(25°~); Ⅵ(26°~); Ⅶ(27°~); Ⅷ(28°~); Ⅸ(29°~)。A(500~); B(1 000~); C(1 500~); D(2 000~); E(2 500~); F(3 000~)。 图 1 澜沧江流域居民区蚊类物种丰富度沿纬度和海拔梯度的变化趋势 Figure 1 Pattern of species richness of mosquitoes along a latitudinal and altitudinal gradient in the Lancang river of Yunnan |
| |
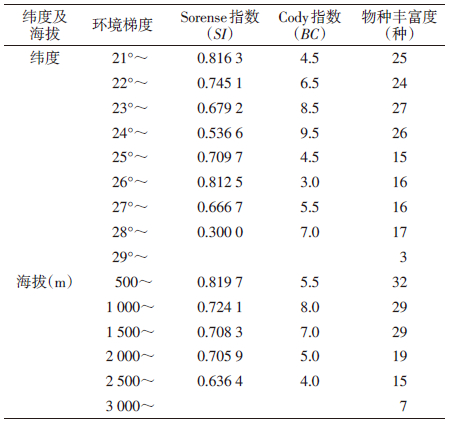 |
蚊类β-多样性(Cody指数)随纬度的增加呈先升高后下降而后再升高的分布格局,高峰于23°~25° N梯度带,为热带(南亚热带)向中亚热带气候、北亚热带向暖温带气候的过渡地带。此区域蚊类物种丰富度也相对较高。反映出由于地理和气候环境的改变,蚊类区系、组成及分布均发生了明显更替。也说明蚊类β-多样性的水平分布格局与地理环境和气候条件的变化有关。
蚊类β-多样性(Cody指数)沿海拔梯度的增加呈先升高后逐步降低的趋势(表 3),最高点位于1 000~2 000 m,山地蚊类区系、物种的组成和丰富度由低山(南亚热气候带)向中山(暖温气候带)变化。蚊类β-多样性指数与相似性系数呈负相关关系,即Cody指数随纬度和海拔梯度的增加,物种相似性系数降低,反之亦然。
2.4 蚊类空间分布的聚类分析与区系分异蚊种沿着纬度梯度水平分布的相似性聚类结果显示为3个主要地域区系类型(图 2a):Ⅰ~Ⅳ带(21°~24° N),应属于东洋区系,水资源丰富,物种多样性最高,代表蚊种有微小按蚊 (An. minimus)、棋斑按蚊(An. tessellatus)和迷走按蚊(An. vagus)等;Ⅴ~Ⅷ带(25°~28° N),地处中亚热带、北亚热带至暖温带区间,属于东洋区系与古北区系的交错带或过渡区,该区域主要为产稻区,蚊类以稻田型孳生三带喙库蚊和中华按蚊为优势种,其密度较高,而物种丰富度相对较低;Ⅸ带(29°~30° N)地处青藏高原的南缘,基本属寒温带气候,属于古北区系,蚊种的组成较为简单,多样性较低,代表种有银带脉毛蚊(Cu. niveitaeniata)和帕氏按蚊(An. pattoni)。

|
| 注:a.Ⅰ(21°~); Ⅱ(22°~); Ⅲ(23°~); Ⅳ(24°~); Ⅴ(25°~); Ⅵ(26°~); Ⅶ(27°~); Ⅷ(28°~); Ⅸ(29°~)。b. A(500~); B(1 000~); C(1 500~); D(2 000~); E(2 500~); F(3 000~)。 图 2 云南省澜沧江流域居民区蚊类沿纬度(a)和海拔(b)梯度聚类分析结果 Figure 2 Cluster analysis of mosquitoes along the latitude and altitude gradient in the Lancang river of Yunnan province |
| |
蚊类物种沿着海拔梯度的垂直分布相似性的聚类结果呈现与纬度梯度(水平分布)近似的特征,划分为3类(图 2b):地处海拔较低的南亚热带气候带A~C带(500~2 000 m),为东洋区系; D~E(2 000~3 000 m)梯度带,D、E带为中低山暖温性气候向亚高山温凉性气候的过渡地带,应为东洋区系向古北区系过渡和交汇地带;地处亚高山地带F(3 000~3 500 m)梯度带,属于寒温带气候类型,蚊类属于古北区系。各区系蚊类代表种与纬度梯度分布的情况基本类似。
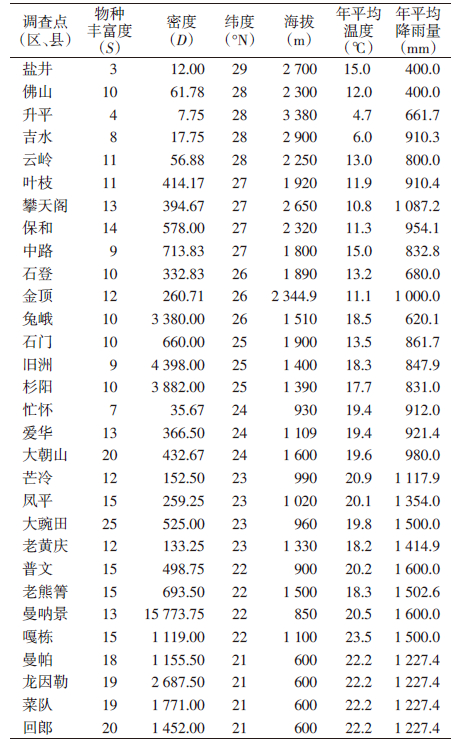
2.5 蚊类物种丰富度、密度与环境因素的关系澜沧江流域12个县(市)30个乡镇蚊类物种丰富度、密度和重要环境因素的关系见表 4。蚊类物种丰富度与纬度、海拔、温度、降雨量和密度均有相关性(r=-0.720、-0.645、0.598、0.675、0.785,均P<0.001),差异有统计学意义;但蚊类密度与纬度、海拔、温度和降雨量差异无统计学意义(r=0.139、0.097、0.164、0.129,均P>0.05),见图 3。蚊类物种丰富度与纬度和海拔呈负相关,与温度和降雨量呈正相关。
 |

|
| 图 3 澜沧江流域居民区蚊类物种丰富度、密度分别与纬度、海拔、温度和降雨量的关系散点图 Figure 3 Scatter plot of relationship between mosquito species richness and density with latitude, altitude, temperature and rainfall in Lancang river |
| |
由于地理和气候条件的影响,云南省澜沧江流域居民区蚊类γ-多样性的空间分布格局总体呈现随纬度和(或)海拔的增高而递减的趋势,随温度和(或)降雨量的增加而逐步增高的趋势。在地理因素中纬度对蚊类多样性的影响较为明显,气候因素中降雨量对蚊类多样性有重要作用。
3 讨 论 3.1 蚊类γ-多样性空间梯度的格局与特征物种多样性随纬度降低而增加是被大多数陆生植物和动物可适应的分布格局[19]。澜沧江流域由北向南贯穿云南省西部,流经区域由于地理位置不同和高度差,从南向北或由低到高依次可出现低山、中山、亚高山、高山、极高山,或热带、亚热带、温带和寒带不同的气候带和森林植被景观。蚊类γ-多样性调查结果显示,<24°纬度的区间物种丰富度较高(24~27种),海拔<2 000 m区间物种丰富度较高(29~32种)。蚊类γ-多样性梯度格局从北向南,由高到低,随着纬度和海拔梯度的下降,气温和降雨量的上升,总体也呈现了由低到高的变化趋势与特征。
此结果与同区域的横断山区(21°~28° N,海拔600~4 500 m)蚤类物种多样性空间分布格局显然不同[20-23]。主要是蚊类幼虫期主要孳生于各类露天的自然水体,而蚤类主要孳生于不同的宿主动物洞穴及窝巢。在同一区域地理因素中,蚊类受纬度影响较大,气候因素中受降雨量的影响较大,山地的雨量分布特征和水湿条件影响蚤类多样性地理分布趋势[21];蚊类以河谷带的中低山地居民区为主,而蚤类以低山至高山的不同森林植被带为主[9]。此外,还可能与研究区地域、尺度大小、海拔高度、气候环境条件、调查和取样方法及研究对象有关[10]。
蚊种是否吸血、活动时间和环境选择等习性不同,但其成蚊一般可分为以夜间或白天活动为主的两大类型。目前还没有较好或通行的,可在不同栖境、昼夜兼顾的调查方法。云南省澜沧江流域应用多种方法调查和记录的蚊类,初步统计达13属约110种[12-17]。本研究利用灯诱捕法对澜沧江流域居民区进行调查发现蚊类7属46种,其中按蚊属16种,库蚊属15种,伊蚊属11种,其他4属各1种。与已记录的13属107种中主要属、种数比较,13属(13:7)、按蚊30种(30:16)、库蚊24种(24:15)和伊蚊24种(24:11),总蚊种110种(110:46),总体数量未达到该区域已知蚊类的一半。说明本次调查结果仅反映和代表云南省西部澜沧江流域居民区蚊类由北向南沿纬度梯度的物种组成和空间分布特征。同时,应进一步增加不同地理气候环境、取样方法的研究,如对不同孳生环境幼虫的捕捉,白天嗜血活动蚊类的诱捕及山地不同森林垂直带蚊类的调查等。
3.2 澜沧江流域蚊类的区系特征我国西南部横断山区独特的地理环境使当地生物两大区系及其分布形成交叉重叠的特征,该地域的区系划界问题存在争议。本研究根据云南省西部澜沧江流域山地居民区各纬度带或海拔带蚊类组成的异同,应用系统聚类分析方法将其划分为东洋区系、两区系交汇或过渡区和古北区系3个主要地域区系类型,与横断山区蚤类区系的空间分布格局及特征[20-23]基本一致,反映了我国西南部横断山区昆虫区系空间分布格局的一般规律和区域特征。
3.3 蚊类γ-多样性空间分布格局与地理、气候因素的关系物种多样性大空间尺度格局与环境因子密切相关,不同自然区域中各物理因子作用不同。澜沧江源于青藏高原,地处滇西横断山区,属青藏高原的南延部分。该区域地势北高南低,气温和雨量由北到南或由高到低有一定的变化规律,该流域整体属于湿润和半湿润地区,但下游雨量充沛而上游雨量较少。不同气候带蚊类组成和分布存在明显差异,说明蚊类多样性分布格局与地理、气候条件的变化有关。蚊类多样性空间分布格局与纬度、海拔两变量呈负相关(r=-0.720、-0.645,均P<0.001),与温度和降雨量呈正相关(r=0.598、0.675,均P<0.001)。纬度和降雨量对澜沧江流域蚊类物种多样性的空间分布格局影响较大。由于地理和气候条件等综合因素的影响,澜沧江流域蚊类γ-多样性的梯度格局总体呈现随纬度和海拔增高而递减的趋势,随气温和降雨量增加而逐步增高的趋势。
但蚊类密度与纬度、海拔、温度和降雨量的相关性较差(r=0.139、0.097、0.164、0.129)。分析发现,在调查区蚊类密度较高的兔峨乡、旧洲村、杉阳镇、曼呐景等(表 3),其居民区周边环境及栖息地以种植水稻为主的河谷台地或坝区、以稻田为主要孳生地的三带喙库蚊物种多度占绝对优势,导致总蚊密度很高(表 1)。说明不同环境取样点,居民区周边的主要栖息地(孳生地)类型对蚊类密度有重要影响,与云南省丽江市研究相似[24]。
云南省西部横断山区地理环境复杂多样,由于不同江河流域所处地理位置、雨量分布特征、气候环境条件的差异,各流域之间蚊类物种多样性的格局与特征亦存在一定差异。因此,不同经、纬度各流域之间蚊类多样性格局形成的云南省区域特征和变化趋势、形成机制与影响因素的关系,仍值得进一步研究。
致谢: 现场调查工作中得到西藏自治区芒康,云南省迪庆、德钦、维西、兰坪、云龙、永平、云县、景谷、宁洱、西双版纳、景洪、勐腊等州(县、市)疾病预防控制中心及各地乡镇卫生院的支持与帮助,特此志谢| [1] | 张荣祖, 林永烈. 中国及其邻近地区兽类分布的趋势[J]. 动物学报,1985,31 (2) :187–197. |
| [2] | 张荣祖. 中国动物地理[M]. 北京: 科学出版社, 1999 : 1 -121. |
| [3] | Rosenzweig ML. Species diversity in space and time[M]. Cambridge: Cambridge University Press, 1995 : 1 -460. |
| [4] | Colwell RK, Lees DC. The mid-domain effect: geometric constraints on the geography of species richness[J]. Trends Ecol Evol, 2000, 15 (2) : 70–76 . |
| [5] | 胡军华, 胡慧建, 蒋志刚. 大空间尺度上物种多样性的分布规律[J]. 应用与环境生物学报,2007,13 (5) :731–735. |
| [6] | 吴永杰, 雷富民. 物种丰富度垂直分布格局及影响机制[J]. 动物学杂志,2013,48 (5) :797–807. |
| [7] | Lyons SK, Willig MR. A hemispheric assessment of scale dependence in latitudinal gradients of species richness[J]. Ecology, 1999, 80 (8) : 2483–2491 . |
| [8] | 龚正达, 吴厚永, 段兴德, 等. 云南横断山区小型兽类物种多样性与地理分布趋势[J]. 生物多样性,2001,9 (1) :73–79. |
| [9] | 葛军旗, 孙肖红, 龚正达, 等. "三江并流"自然遗产地澜沧江流域居民区蚊类多样性的空间分布格局[J]. 生物多样性,2008,16 (1) :24–33. |
| [10] | 张菊仙, 龚正达, 李四全, 等. "三江并流"自然遗产地怒江流域居民区蚊类多样性的空间分布格局[J]. 寄生虫与医学昆虫学报,2008,15 (4) :213–222. |
| [11] | 李栋, 龚正达, 李四全, 等. "三江并流"自然遗产地金沙江流域居民区蚊类物种多样性与多度空间分布格局及其关系[J]. 寄生虫与医学昆虫学报,2011,18 (4) :204–215. |
| [12] | 张云智, 张海林, 龚正达, 等. 云南省澜沧江下游地区蚊虫群落及地理生态位的研究[J]. 地方病通报,2001,16 (1) :74–80. |
| [13] | 周红宁, 杜尊伟, 肖育江, 等. 云南省勐腊县人房蚊虫群落特征研究[J]. 中国媒介生物学及控制杂志,2002,13 (3) :181–182. |
| [14] | 王学忠, 杜尊伟, 卢勇荣, 等. 云南南部野外与村内蚊虫群落特征的比较研究[J]. 中国媒介生物学及控制杂志,2003,14 (5) :348–350. |
| [15] | 王学忠, 杜尊伟, 李菊昇, 等. 不同生境蚊类生态学研究[J]. 中国媒介生物学及控制杂志,2005,16 (2) :98–100. |
| [16] | 李佳, 周红宁, 王丕玉, 等. 澜沧江中下游地区居民点蚊虫多样性的初步研究[J]. 中国媒介生物学及控制杂志,2009,20 (2) :104–107. |
| [17] | 王丕玉, 周红宁, 吴超, 等. 澜沧江下游地区蚊虫种类、分布及其孳生习性调查[J]. 中国热带医学,2011,11 (6) :657–659. |
| [18] | 赵志模, 郭依泉. 群落生态学原理与方法[M]. 重庆: 科学技术文献出版社重庆分社, 1990 : 1 -288. |
| [19] | Hillebrand H. On the generality of the latitudinal diversity gradient[J]. Am Nat, 2004, 163 (2) : 192–211 . |
| [20] | 龚正达, 吴厚永, 段兴德, 等. 云南临沧地区蚤类群落生态学与区系研究[J]. 寄生虫与医学昆虫学报,2000,7 (3) :160–169. |
| [21] | 龚正达, 吴厚永, 段兴德, 等. 云南横断山区蚤类物种多样性的地理分布趋势与重要环境因素的关系[J]. 生物多样性,2001,9 (4) :319–328. |
| [22] | 龚正达, 吴厚永, 段兴德, 等. 云南横断山区蚤类物种丰富度与区系的垂直分布格局[J]. 生物多样性,2005,13 (4) :279–289. |
| [23] | 龚正达, 张丽云, 段兴德, 等. 中国"三江并流"纵谷地蚤类丰富度与区系沿纬度梯度的水平分布格局[J]. 生物多样性,2007,15 (1) :61–69. |
| [24] | 连宏宇, 臧颖惠, 边长玲, 等. 云南丽江市自然风景区居民点蚊类多样性研究[J]. 寄生虫与医学昆虫学报,2010,17 (4) :212–217. |