 2016, Vol. 27
2016, Vol. 27扩展功能
文章信息
- 仝振东, 金永富, 易井萍, 刘洁楠
- TONG Zhen-dong, JIN Yong-fu, YI Jing-ping, LIU Jie-nan
- 舟山市海岛地区蚊媒种群及其蚊媒传染病病原体分布研究
- Study on distribution of mosquito vectors and associated communicable disease pathogens in islands of Zhoushan, China
- 中国媒介生物学及控制杂志, 2016, 27(2): 168-171
- Chin J Vector Biol & Control, 2016, 27(2): 168-171
- 10.11853/j.issn.1003.8280.2016.01.019
-
文章历史
- 收稿日期: 2015-12-14



舟山市位于浙江省东部,121°30′~123°25′E,29°32′~31°04′N之间,由1 390个岛屿组成。2011年6月30日,国务院正式批准设立浙江省舟山群岛新区,随着舟山群岛新区的建设推进,大宗商品国际物流基地和综合保税港区迅速发展,舟山市病媒生物生态环境可能发生改变,病媒生物的输入风险以及登革热等蚊媒传染性疾病的输入风险逐渐增加。
我国蚊类至少有3亚科21属52亚属395种(亚种)[1],是昆虫纲中分布范围最广的医学昆虫之一,其可传播登革热、基孔肯雅热、流行性乙型脑炎(乙脑)等虫媒传染病,为传播病原体种类最多的媒介生物。为了解舟山地区蚊虫的种群分布及病原体携带状况,评估舟山地区发生蚊媒传染病的暴发风险,本研究调查了舟山市不同地区的蚊虫种群分布,并检测相关病原体,为该地区蚊媒传染病的防控提供科学依据。
1 材料与方法 1.1 材料MobileMapperTM 6.0 GPS(全球定位系统)定位仪,40目标准铜筛,多功能便携式pH计,博迅BOXUN SPX-300IC人工气候箱,EZ4D体视显微镜,功夫小帅诱蚊灯,乙醚。
1.2 幼蚊采集采用勺舀法(Dipper,500 ml),参照Silver《Mosquito ecology-field sampling methods》,以普陀区、定海区、临城新区等为中心,辐射周边若干个岛屿及其乡镇街道,对辖区内河流、湖泊、池塘、农田水沟等水体按“Z”形路线进行随机抽样,同时利用GPS定位仪对靶标位点进行标记。每批水样容积约为50 ml,去除杂质,单独置于250 ml透明塑料瓶中,同时测量水体pH值。将野外采集到的蚊幼虫带回实验室进行人工饲养,温度(25±1)℃,湿度50%~70%,光照时间L∶D=12 h∶12 h。待成蚊羽化后,分类鉴定。
1.3 成蚊采集采用光诱捕法,按东、南、西、北、中5个方位,选择牲畜养殖场、滩涂度假点、公园、大型停车场、花卉苗圃等易于蚊虫孳生场所,每处悬挂5盏功夫小帅诱蚊灯,18:00-21:00布放。将捕获成蚊用乙醚麻醉后置于2 ml冻存管中,标记后带回实验室分类鉴定并计数,同时记录当天气温、湿度、海拔高度、经纬度(GPS定位标记)等相关信息。
1.4 病原体检测根据S--nchez-Seco等[2, 3, 4, 5]提供的6对通用简并引物(表 1),利用多重PCR和巢式PCR对白纹伊蚊(Aedes albopictus)、东乡伊蚊(Ae. togoi)、中华按蚊(Anopheles sinensis)等携带病毒率进行初筛,检测其基孔肯雅病毒(CHIKV)、登革热病毒(DENV)、乙脑病毒(JEV)等携带状况。

对蚊虫采集结果和实验室检测结果进行描述性分析,计算优势种指数、雌雄比等。
2 结 果 2.1 幼蚊监测对普陀山、朱家尖、六横岛、金塘岛、定海以及临城等20多个乡镇及街道的酒坛、小塑料桶、水缸、轮胎、罐头瓶、竹桩、油漆桶、海边岩石坑和泡沫箱等积水容器进行调查,覆盖范围29°43′31″~30°15′57″N,122°6′31″~122°15′58″E,海拔105~358 m,属于低海拔区域。共采集蚊幼虫样本106份,经实验室人工气候箱孵育培养后〔(25±1)℃、RH 50%~60%〕,羽化后收获成蚊1 358只。经鉴定,共包括2亚科5属11种(表 2),其中库蚊属环带库蚊(Culex annulus)和杵蚊属蛛形杵蚊(Tripteroides aranoides)为舟山市新发现种。

登革热和基孔肯雅热的传播媒介白纹伊蚊为当地的主要优势种,羽化数为641只,指数为0.472,孳生的水体环境为弱碱性。其次为乙脑及马来丝虫病传播媒介东乡伊蚊,羽化数为384只,指数为0.283,除个别地域外,其分布仅限于沿海一带的临时性岩石积水中,幼虫能生活在高盐水的水体中(平均pH值为8.13)。致倦库蚊(Cx. pipiens quinquefasciatus)采集量虽然有限,但其成蚊的羽化率高。环带库蚊、迷走库蚊(Cx. vagans)和蛛形杵蚊种群密度极低,且其繁殖对于生境的要求比较高。而褐尾库蚊(Cx. fuscanus)和三带喙库蚊(Cx. tritaeniorhynchus)却恰恰相反,易孳生于含氮高的水体,前者主要孳生于露天粪坑,后者多孳生在养殖场附近的农田水沟中。
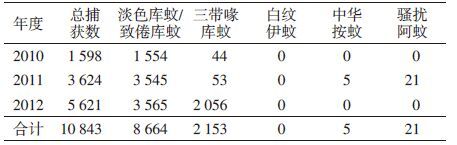
2.2 成蚊监测2010-2012年舟山地区共捕获成蚊10 843只,以淡色库蚊/致倦库蚊(Cx. pipiens pallens)为主,占捕蚊总数的79.90%(8 664/10 843)(表 3),为优势蚊种;其次是三带喙库蚊,占捕蚊总数的19.86%(2 153/10 843);其他蚊种如中华按蚊、骚扰阿蚊(Armigeres subalbatus)等捕获量均较少,未捕到白纹伊蚊。
 |
捕获的白纹伊蚊、东乡伊蚊、淡色库蚊/致倦库蚊、三带喙库蚊、骚扰阿蚊等成蚊均未检测到CHIKV、DENV和JEV,结果全部为阴性。
3 讨 论2009年舟山跨海大桥正式开通以及2011年国务院批复成立浙江省舟山群岛新区,宣告舟山市进入了一个全新的发展时代。另外,作为一个旅游资源极为富饶的海岛城市,有“海天佛国”普陀山、“沙雕故乡”朱家尖等风景名胜区,每年吸引着600多万海内外游客并以15%的速度逐年递增。随着舟山贸易和旅游的发展,人员往来频繁,登革热、基孔肯雅热、疟疾等蚊媒传染病的输入风险增加,而且舟山地区气候适合蚊虫生长繁殖,一旦有病例输入有发生暴发的风险,同省的慈溪和义乌市先后曾发生输入病例并引起本地登革热暴发[6, 7]。 幼虫监测结果显示舟山市优势种指数最高的依次为白纹伊蚊、东乡伊蚊和致倦库蚊,它们分别是登革热、基孔肯雅热、乙脑等多种传染病的传播媒介,提示舟山地区具备该类病毒传播条件。但影响幼虫调查因素诸多,关键在于采样点的类型或生境,由于不同蚊种的适生生境不同,在清洁的积水容器、臭水沟和稻田等大水体采样所得到的监测结果会截然不同。
成蚊监测结果显示舟山市优势蚊种为致倦库蚊和三带喙库蚊,此与监测方法有关,因伊蚊是日间活动,且对光的敏感性较弱,所以在18:00-21:00用光谱诱蚊灯很难捕获伊蚊。王采典等[8]报道致倦库蚊为舟山口岸的优势蚊种,傅桂明等[9]报道致倦库蚊和三带喙库蚊在浙江省广泛分布,与本研究结果类似。此外,与舟山市距离较近的上海市常见蚊种也为致倦库蚊和三带喙库蚊[10]。
幼虫监测法和成蚊监测法各有利弊。幼虫监测受采样点的影响较大,不同的生境其优势蚊虫不同。成蚊监测受监测器械和时间的影响,诱蚊灯对白纹伊蚊的诱捕效果有限。本研究结合幼蚊监测和成蚊监测的优点,掌握了舟山地区的优势蚊种,为蚊媒传染病的预防控制奠定了基础。 然而,本研究在所捕获的蚊虫中未检测出DENV、CHIKV和JEV,可能由于舟山地区非登革热、基孔肯雅热和乙脑的疫源地。蚊虫中未携带病毒,可能由于样本采集、样本保存以及检测方法局限等。慈溪和义乌市登革热暴发时均未从蚊虫中检测出DENV[6, 7],也说明检测蚊虫中病毒难度较高。
综上所述,通过掌握舟山地区的蚊虫种群分布,结合幼虫和成虫监测,白纹伊蚊、东乡伊蚊、致倦库蚊和三带喙库蚊为舟山市的优势蚊种。尽管未检测出靶标病毒,一旦有病例输入,本地也有暴发的风险,需要加强蚊媒传染病防控。
| [1] | 瞿逢伊. 我国蚊虫种质资源现状及其共享利用[J]. 中国寄生虫学与寄生虫病杂志,2006,24增刊:13-16. |
| [2] | Sánchez-Seco MP,Rosario D,Quiroz E,et al. A generic nested-RT-PCR followed by sequencing for detection and identification of members of the alphavirus genus[J]. J Virol Methods, 2001,95(1/2):153-161. |
| [3] | Sánchez-Seco MP,Echevarria JM,Hernández L,et al. Detection and identification of Toscana and other phleboviruses by RT-nested-PCR assays with degenerated primers[J]. J Med Virol, 2003,71(1):140-149. |
| [4] | Sánchez-Seco MP,Rosario D,Domingo C,et al. Generic RT-nested-PCR for detection of flaviviruses using degenerated primers and internal control followed by sequencing for specific identification[J]. J Virol Methods,2005,126(1/2):101-109. |
| [5] | Sánchez-Seco MP,Rosario D,Hernández L,et al. Detection and subtyping of dengue 1-4 and yellow fever viruses by means of a multiplex Rt-nested-PCR using degenerated primers[J]. Trop Med Int Health,2006,11(9):1432-1441. |
| [6] | Yang TC,Lu L,Fu GM,et al. Epidemiology and vector efficiency during a dengue fever outbreak in Cixi,Zhejiang province,China[J]. J Vector Ecol,2009,34(1):148-154. |
| [7] | Sun JM,Lin JF,Yan JY,et al. Dengue virus serotype 3 subtype Ⅲ, Zhejiang province, China[J]. Emerg Infect Dis,2011,17(2):321-323. |
| [8] | 王采典,马卫星,秦伟平. 舟山口岸蚊类种群及季节消长调查[J]. 中国国境卫生检疫杂志,2003,26(6):345-347. |
| [9] | 傅桂明,龚震宇,杨天赐,等. 2009年浙江省诱蚊灯法监测蚊虫结果分析[J]. 中国媒介生物学及控制杂志,2010,21(5):422-423. |
| [10] | 周正斌,吕山,张仪,等. 上海市蚊媒种类、分布及其病原[J]. 中国媒介生物学及控制杂志,2015,26(1):28-32. |