 2016, Vol. 27
2016, Vol. 27扩展功能
文章信息
- 刘正祥, 高子厚, 尹家祥, 杜春红, 洪梅, 李玉琼, 宋志忠
- LIU Zheng-xiang, GAO Zi-hou, YIN Jia-xiang, DU Chun-hong, HONG Mei, LI Yu-qiong, SONG Zhi-zhong
- 云南省居民区室内小型兽类体表蚤类与宿主关系研究
- Study on the relationship between fleas and indoor small mammal hosts in residential areas in Yunnan province
- 中国媒介生物学及控制杂志, 2016, 27(2): 151-154
- Chin J Vector Biol & Control, 2016, 27(2): 151-154
- 10.11853/j.issn.1003.8280.2016.02.015
-
文章历史
- 收稿日期: 2015-11-10



2 大理大学公共卫生学院, 云南大理 671000
2 Dali University, School of Public Health
云南省位于我国西南边陲,多山地形,地势西北高、东南低,以山地高原为主。由于地理位置特殊、低纬度高原,地形地貌复杂,兼具低纬气候、季风气候及山地高原气候特点。因得天独厚的自然条件,境内动植物种类丰富,素有“动植物王国”之称。在动物地理区划上,隶属于东洋界中印亚界西南区的西南山地亚区和华南区的滇南山地亚区[1]。蚤类作为一类重要医学昆虫,主要寄生于哺乳动物和鸟类体表,为鼠疫和鼠源性斑疹伤寒等疾病的传播媒介[2]。居民区作为一个特殊的动物栖息生境,受社会发展和人为活动影响较大,动物及其寄生蚤群落相对于森林区、农耕地等生境较为简单,其物种丰富度和多样性等群落指标与森林型等自然群落明显不同[3],而居民区室(院)内环境更为特殊,为人类活动的中心区域,也存在畜、禽类。按解宝琦和曾静凡[4]1992年根据中国地理区划划分的云南省5个动物地理小区,本研究于2009-2013年对居民区室内蚤类及其宿主动物进行了抽样调查,以了解云南省不同动物地理小区居民区室内蚤类及其宿主动物的分布情况,并运用生态位宽度和物种多样性指数等指标,研究不同动物地理小区的蚤类与宿主动物的关系。
1 材料与方法 1.1 研究样区根据云南省5个不同动物地理区域面积大小,在每区域内选取≥6个县(市、区),共43个县(市、区)作为调查样区。主要分类:Ⅰ. 横断山中部小区:剑川、云龙、兰坪、保山市隆阳区、玉龙、香格里拉和德钦7个县(市、区);Ⅱ. 横断山南部小区:云县、凤庆、耿马、景谷、宁洱、施甸、大理、弥渡8个县(市、区);Ⅲ. 滇东高原小区:祥云、永仁、南华、弥勒、石屏、开远、丘北、砚山、广南、罗平、新平、昭通市昭阳区、昆明和宜良14个县(市、区);Ⅳ. 滇西高原小区:腾冲、龙陵、梁河、盈江、瑞丽、芒市和陇川7个县(市、区);Ⅴ. 滇南山地小区:勐海、勐腊、景洪、孟连、马关、江城和永德7个县(市、区)。其中,剑川、景谷、大理、罗平、昭通市昭阳区、砚山、瑞丽、孟连、江城等县(市、区),采用了2010-2013年间调查方法相同的鼠疫宿主及媒介监测数据。
1.2 调查方法每个县(市、区)抽取4个自然村,以自然村或街道为采样地点,每个自然村随机抽取20个住户进行调查。调查时,在各住户庭院围墙以内,包括客厅、卧室、厨房、畜圈、仓库、过道及庭院内绿化及菜园地等处放置工具。无围墙的住户仅在室内布放工具。每户放置鼠笼5个,以鲜炸油条或火腿肠为诱饵,连续放置3夜。每日早晨检查鼠笼,如捕到鼠,则更换新笼并继续放置于同一位置。捕获鼠类标本一鼠一袋密封,带回实验室麻醉后鉴定,分类编号记录,按常规方法收集体表全部寄生蚤,并将蚤类制成玻片标本,在光学显微镜下鉴定到种。
1.3 统计学方法[5]物种丰富度(S):S为某环境内物种数;
生态位宽度(Bi):采用Levins模型,Bi=1/S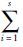 Pin2,其中,Pin2为在资源序列上,寄生蚤i利用宿主动物第n级资源的比例;
Pin2,其中,Pin2为在资源序列上,寄生蚤i利用宿主动物第n级资源的比例;
多样性指数(H′):采用Shannon-Wienner多样性公式:H′=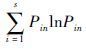 ;
相关系数(r):采用直线回归。
;
相关系数(r):采用直线回归。
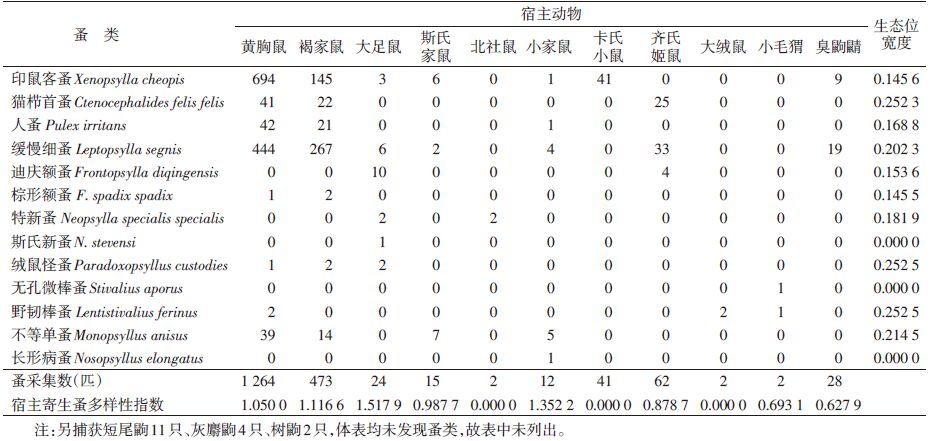
在5个不同地理小区43个县(市、区)172个村(样点),共捕获鼠类宿主动物1 582只,分属3目5科10属14种,并获体表寄生蚤1 925匹,分属5科11属13种,见表 1。其中Ⅰ小区捕获宿主动物10种188只,黄胸鼠(Rattus tanezumi)捕获量最多,占该区捕获总数的37.23%(70/188),其次是大足鼠(R. nitidus)(68/188,36.17%)和小家鼠(Mus musculus)(14/188,7.45%);捕获蚤8种311匹,其中缓慢细蚤(Leptopsylla segnis)最多,占57.88%(180/311),其次为印鼠客蚤(Xenopsylla cheopis)(58/311,18.65%)和猫栉首蚤(Ctenocephalides felis felis)(35/311,11.25%)。Ⅱ小区捕获宿主动物5种246只,其中黄胸鼠最多,占该区捕获总数的82.93%(204/246),其次是褐家鼠(R. norvegicus),占13.41%(33/313),其他鼠数量较少;捕获蚤4种313匹,印鼠客蚤最多,占77.32%(242/313),其次是缓慢细蚤(53/313,16.93%)和不等单蚤(Monopsyllus anisus)(15/313,4.79%)。Ⅲ小区捕获宿主动物7种553只,其中褐家鼠最多,占该区捕获总数的43.40%(240/553),其次是黄胸鼠(163/553,29.48%)和小家鼠(123/553,22.24%);获蚤8种615匹,缓慢细蚤最多,占53.33%(328/615),其次是印鼠客蚤(199/615,32.36%)和人蚤(Pulex irritans)(44/615,7.15%)。Ⅳ小区捕获宿主动物9种310只,其中黄胸鼠捕获量最多,占该区捕获总数的89.03%(276/310),其次是臭鼩鼱(Suncus murinus)(19/310,6.13%)和大足鼠(4/310,1.29%);获蚤7种372匹,印鼠客蚤最多,占80.91%(301/372),其次是缓慢细蚤(53/372,14.25%)和不等单蚤(8/372,2.15%)。Ⅴ小区捕获宿主动物5种285只,其中黄胸鼠捕获量最多,占该区捕获总数的76.14%(217/285),其次是褐家鼠(52/285,18.25%)和小家鼠(9/285,3.16%);获蚤5种314匹,缓慢细蚤最多,占51.27%(161/314),其次是印鼠客蚤(99/314,31.53%)和猫栉首蚤(30/314,9.55%)。捕获短尾鼩(Anowrosorex squamipesi)11只、灰麝鼩(Crocidura attenuata)4只和树鼩(Tupaia belangeri)2只,体表均未发现蚤。
 |
13种蚤的生态位宽度结果见表 1。其中绒鼠怪蚤(Paradoxopsyllus custodies)、野韧棒蚤(Lentistivalius ferinus)、猫栉首蚤、不等单蚤和缓慢细蚤的寄主生态位宽度较高,分别为0.252 5、0.252 5、0.252 3、0.214 5和0.202 3;斯氏新蚤(Neopsylla stevensi)、无孔微棒蚤(Stivalius aporus)和长形病蚤(Nosopsyllus elongatus)仅寄生于一种宿主动物,表明它们的寄主生态位最窄,为0。
2.3 宿主动物寄生蚤多样性11种宿主动物中,大足鼠寄生蚤多样性指数最高(1.517 9),其次为小家鼠(1.352 2)和褐家鼠(1.116 6)。北社鼠(Niviventer confucianus)、卡氏小鼠(M. caroli)和大绒鼠(Eothenomys miletus)仅有一种寄生蚤,多样性指数为0,说明这几种宿主动物寄生蚤丰富度较低,见表 1。
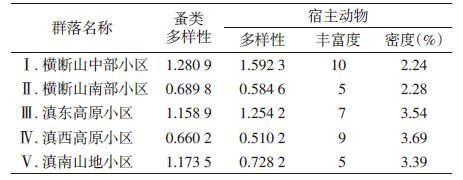
2.4 各动物地理小区蚤类多样性与宿主多样性、丰富度及宿主密度的关系由表 2可见,5个动物地理小区中,Ⅰ小区蚤类多样性指数最高,Ⅴ小区次之,Ⅳ小区最低;宿主动物多样性则Ⅰ小区最高,Ⅲ小区次之,Ⅳ小区最低;宿主动物种类数Ⅰ小区最多,Ⅳ小区和Ⅲ小区次之;宿主动物密度以Ⅳ和Ⅲ小区较高,Ⅰ小区最低。经相关分析,各小区蚤类多样性指数与宿主动物多样性指数、丰富度及宿主密度间差异无统计学意义(r=0.820 4、0.290 7、-0.084,均P>0.05)。
 |
寄生蚤分布与啮齿动物分布环境条件密切相关。某一生境蚤类群落中种的构成和不同种类蚤的丰富度受宿主的种类构成、相对丰富度及生境本身特性的影响[6]。居民区室内作为蚤类宿主动物栖息环境之一,生境条件相对单一,物种丰富度及物种多样性与农田耕地及森林等生境比较相对较低,而物种多样性可较全面地反映某地区的物种组成和分布状况[7]。由表 2可知,云南省不同动物地理小区居民区室内蚤类多样性为Ⅰ>Ⅴ>Ⅲ>Ⅱ>Ⅳ;宿主动物多样性为Ⅰ>Ⅲ>Ⅴ>Ⅱ>Ⅳ。各地理小区居民区室内蚤类以印鼠客蚤和缓慢细蚤为优势种,其中,横断山南部和滇西高原小区印鼠客蚤优势明显,而其他地理小区以缓慢细蚤优势突出。蚤类宿主动物除横断山中部小区以黄胸鼠和大足鼠为优势鼠种外,其他小区均以黄胸鼠和褐家鼠为优势鼠种,其中横断山中部、滇西高原和滇南山地小区黄胸鼠优势最为明显。另外,由于云南省地形多山,除坝区居民区外,还有大面积的山地居民区,其与山地耕地、森林等生境成交错分布,因此,在居民区室内偶尔捕获一些野栖小兽,如树鼩、齐氏姬鼠(Apodemus chevrieri)和大绒鼠等。云南省不同动物地理小区居民区室内环境大同小异,但其地理位置、海拔、地形等不同而形成不同的自然地理生态景观和气候差异,这些因素均可影响宿主动物分布的多样性。同时,景观类型决定了蚤类的种类及物种多样性[1, 4, 7]。
蚤类是全变态昆虫,其生活史可分为卵、幼虫、蛹和成虫4个时期,其中幼虫是非寄生的(除Uropsylla tasmanica外),其以巢穴中的有机物质为生,成虫营寄生生活,即寄生于宿主动物体外,吸取宿主动物血液,与宿主动物形成寄生和被寄生的关系[8, 9]。本研究所获的13种蚤中,印鼠客蚤和缓慢细蚤的寄生宿主最多,可以分别寄生于7种宿主体表,不等单蚤、猫栉首蚤、人蚤、绒鼠怪蚤、野韧棒蚤所寄生的小兽宿主种类也相对较多,宿主范围也较宽。斯氏新蚤、无孔微棒蚤和长形病蚤仅寄生于1种宿主动物,宿主范围较窄(表 1)。从寄生虫经典理论看,蚤类所寄生的宿主范围越宽,其宿主特异性越低,反之亦然。仅以蚤类所寄生的宿主范围判定和比较蚤类的宿主特异性存在一定的误差,而利用生态学中的生态位宽度可克服此不足[10]。生态位宽度可以用来衡量某物种种群在群落内对某资源序列的利用幅度。郭宪国等[11]应用Levins生态位宽度研究革螨与宿主小兽的关系,将其作为评价革螨与宿主特异性的指标,并可间接评价革螨等节肢动物的媒介潜能。相关研究也表明,当以宿主动物为资源序列时,体表寄生虫的生态位宽窄与其宿主动物特异性呈相反的关系,即生态位越宽,所利用的食物资源越多,对宿主选择的特异性越低,该寄生虫的适应性越强,反之亦然[9, 12, 13]。本研究13种蚤的宿主范围与生态位宽度的变化趋势基本一致,但并不完全吻合,表明宿主范围只能大致判定宿主的特异性,而生态位宽度可更好地判定蚤类的宿主特异性。从寄生蚤类的数量方面进行计算,蚤种对宿主动物的选择进行量化分析,同时考虑了某种蚤所寄生的宿主种类以及寄生数量,有助于对不同宿主动物体外寄生蚤种之间比较,能更全面地反映蚤类与宿主动物的关系,避免误差。
调查显示,在云南省居民区室内当蚤类成虫以所寄生的宿主动物为资源序列时,Levins生态位宽度最宽的是绒鼠怪蚤(0.252 5)和野韧棒蚤(0.252 5),表明13种蚤类中,这2种蚤的宿主特异性最低,其次是猫栉首蚤、不等单蚤、缓慢细蚤、特新蚤和人蚤等。斯氏新蚤、无孔微棒蚤和长形病蚤其生态位宽度为0,表明它们的宿主特异性最高。虽然印鼠客蚤可以寄生多种小兽,但主要在黄胸鼠体表,故其宿主特异性较高。然而蚤类对宿主的选择性不仅取决于蚤种本身的特性,也受湿度、温度、海拔、植被、成虫对饥饿的忍受程度、不同动物地理景观以及外界环境变化等因素影响。因此某些蚤种分布地区不同其宿主组成以及宿主的特异性不同,或某些蚤种因长时间脱离宿主或游离于野外地面,从而发生“饥不择食”、偶然寄生的现象[2]。
本研究表明,蚤类多样性与宿主动物多样性、丰富度和密度无相关性。此结果与龚正达等[7]、张云智等[14]的研究结果相似。可能由于蚤类的生存要求和适应能力与宿主动物并不完全一致,分布不如宿主广,其幼虫期自由生活,不仅需要适合成虫期的环境条件,如宿主动物的血液、毛皮质量以及生态习性和微小环境等,还需幼虫期的环境条件,如营养物质、温湿度、发育场所等[2]。因此蚤种类和数量的分布与宿主动物的分布并不完全一致。
蚤类通过吸血活动可作为鼠疫、鼠型斑疹伤寒、野兔热、Q热、假结核菌病及犬复孔绦虫病等人畜共患病的传播媒介,其中传播最严重的是鼠疫[2]。本调查中印鼠客蚤、人蚤、不等单蚤、棕形额蚤和特新蚤等分别是云南省家、野两型鼠疫的媒介蚤[15]。14种宿主动物中除小毛猬以外,其他小兽均为云南省家鼠或野鼠鼠疫疫源地的宿主动物,其中黄胸鼠和齐氏姬鼠是鼠疫主要宿主[16, 17]。目前云南省鼠疫处于静息期,多年未发生鼠疫流行,但居民区室内为人-鼠-蚤之间关系最为密切的场所,仍应加强鼠疫监测及预警工作,提高居民的灭鼠、灭蚤、防病意识,防患于未然。
志谢 参与此项调查工作的还有县(市、区)疾病预防控制中心的相关人员,特此志谢
| [1] | 张荣祖. 中国动物地理[M]. 北京:科学出版社,2011:181-222. |
| [2] | 吴厚永. 中国动物志. 昆虫纲. 蚤目[M]. 2版. 北京:科学出版社,2007:45-71,116-127. |
| [3] | 龚正达,吴厚永,段兴德,等. 云南临沧地区蚤类群落生态学与区系研究[J]. 寄生虫与医学昆虫学报,2000,7(3):160-169. |
| [4] | 解宝琦,曾静凡. 云南蚤类志[M]. 昆明:云南科技出版社,2000:22-41. |
| [5] | 赵志模,郭依泉. 群落生态学原理与方法[M]. 重庆:科学技术文献出版社重庆分社,1990:73-119,147-172. |
| [6] | 杨孔,张建漂,吴梦玲,等. 生境对蚤类群落及其生活史的影响[J]. 中国媒介生物学及控制杂志,2011,22(1):86-88. |
| [7] | 龚正达,吴厚永,段兴德,等. 云南横断山区蚤类物种多样性的地理分布趋势与重要环境因素的关系[J]. 生物多样性, 2001,9(4):319-328. |
| [8] | 郭天宇. 蚤类生物学与鼠疫[J]. 生物学通报,2002,37(1):26-27. |
| [9] | Laudisoit A, Leirs H, Makundi R, et al. Seasonal and habitat dependence of fleas parasitic on small mammals in Tanzania[J]. Integr Zool,2009,4(2):196-212. |
| [10] | 张胜勇,吴滇,郭宪国,等. 云南省19县市小兽体表蚤类的生态位研究[J]. 寄生虫与医学昆虫学报,2008,15(4):233-237. |
| [11] | 郭宪国,叶炳辉,顾以铭,等. 云南西部革螨生态位研究[J]. 寄生虫与医学昆虫学报,1994,1(4):47-53. |
| [12] | 郭天宇,许荣满. 喜马拉雅山南坡蚤类营养生态位的研究[J]. 应用生态学报,1999,10(3):67-70. |
| [13] | 郭天宇,许荣满. 北京东灵山地区鼠类体外寄生虫营养生态位的研究[J]. 中国媒介生物学及控制杂志,2002,13(4):257-259. |
| [14] | 张云智,龚正达,冯锡光,等. 云南白草岭地区蚤类与宿主动物关系的研究[J]. 地方病通报,2002,17(2):22-24. |
| [15] | 龚正达,于心,刘起勇,等. 中国鼠疫自然疫源地分型研究Ⅵ.鼠疫媒介生物学特征[J]. 中华流行病学杂志,2012,33(8):818-822. |
| [16] | 高子厚,董兴齐,宋志忠,等. 云南省自然感染鼠疫菌宿主动物的研究[J]. 地方病通报,2008,23(3):15-18,24. |
| [17] | 秦长育,许磊,张荣祖,等. 中国鼠疫自然疫源地分型研究Ⅴ.鼠疫宿主生物学特征[J]. 中华流行病学杂志,2012,33(7):692-697. |