 2016, Vol. 27
2016, Vol. 27扩展功能
文章信息
- 黄涛华, 刘霖, 杜春红, 洪梅, 刘正祥, 高子厚
- HUANG Tao-hua, LIU Lin, DU Chun-hong, HONG Mei, LIU Zheng-xiang, GAO Zi-hou
- 滇西南不同景观小型兽类及其媒介分布与鼠疫关系
- The relationship between the distribution of small mammals, vectors and plague in different landscapes in Southwest Yunnan
- 中国媒介生物学及控制杂志, 2016, 27(2): 112-116
- Chin J Vector Biol & Control, 2016, 27(2): 112-116
- 10.11853/j.issn.1003.8280.2016.02.005
-
文章历史
- 收稿日期: 2015-10-10




2 大理大学公共卫生学院, 云南大理 671000
2 School of Public Health, Dali University
滇西南地区位于云南省横断山脉南延部,海拔400~2 100 m,属于南亚热带季风区和北热带湿润区,光照充足,降雨丰富,境内山势趋于缓和,河谷开阔,山间盆地较大而多,山区森林植被为常绿阔叶林和热带雨林,是云南省家鼠鼠疫疫源地的重要组成部分。小型兽类(主要包括啮齿类、食虫类、兔形类、翼手类及部分小型食肉类动物)及寄生蚤类是鼠疫的重要宿主动物和传播媒介,是维持鼠疫自然疫源性和鼠疫流行的重要条件[1]。多数研究集中在局部地区宿主、媒介分类、种群结构、密度调查、地理分布及季节变化等[2, 3, 4, 5],动物鼠疫监测仍以宿主、媒介的密度和构成为主要指标,尚未从景观流行病学角度系统研究不同景观类型差异对鼠疫发生频率和分布规律的综合影响。本研究以滇西南6个县为调查的空间范围,调查滇西南家鼠鼠疫疫源地不同景观生态系统小兽及寄生蚤群落结构、物种多样性及其空间分布格局,进一步了解其空间分布特征与鼠疫流行的关系。
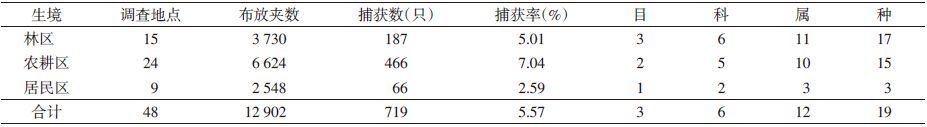
1 材料与方法 1.1 调查范围和时间2011-2012年旱季在云县、永德、景谷、宁洱、勐海、勐腊6个县,对城镇居民点景观(居民区)、农田景观(农耕区)及自然与自然保护景观(林区)3类生态系统中对不同小兽及寄生蚤调查取样。
1.2 调查方法 1.2.1 材料收集对3类景观生态系统林区选取15个调查地点,农耕区24个调查地点,林区和农耕区按夹线法布放鼠夹,每样点置夹≥200个,每25个排成一直线,间距5 m,行距20 m,林区共布放3 730夹次,农耕区共布放6 624夹次。次晨用密封鼠袋收集小兽标本,带回实验室麻醉后按常规方法收集体表的全部蚤类置70%乙醇保存。居民区以自然村或街道为最小单位,选取9个调查地点,按笼日法选择10~20户,每户布放5~10个鼠笼,连续置笼3昼夜;每调查点布放鼠笼≥300个,样本采集同上。
1.2.2 小兽鉴定应用形态学方法,对捕获的小兽先进行性别鉴定和形态特征测量,测量工具包括卷尺、电子秤、标签、记录本等。再依据各种小兽的外部特征及其在云南省分布区域进行现场物种鉴定,对外形相似或少数难以鉴定的种类,制作头骨皮毛标本,以头骨、牙齿等特征进一步鉴定种类。
1.2.3 蚤类鉴定蚤类标本经30%、50%、70%、90%和95%乙醇逐级脱水后,以乙醇/二甲苯(1∶1)、二甲苯透明后用冷杉胶封制玻片标本,应用形态学方法,在显微镜下对所制标本鉴定种类。
1.3 数据统计分析 1.3.1 构成比、染蚤率、蚤指数计算[6]
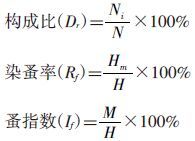
用Shannon-Winner指数公式: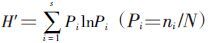 分别计算各景观小兽及其寄生蚤群落物种多样性指数( 式中,H′为多样性指数;Pi为第i种的个体比例);以Pielou(1949) 的公式J=H′/lnS计算小兽及其寄生蚤群落的均匀度指数(式中,J为均匀度,S为种类数);Simpson(1949) 的公式C′=∑(ni/N)2计算各群落的生态优势度(式中C′为生态优势度,ni为每一种的个体数,N为总个体数);Jaccard相似性系数公式(CJ)=j/(a+b-j),式中,j为两种景观系统共有种数,a和b分别为景观A和景观B的物种数。
分别计算各景观小兽及其寄生蚤群落物种多样性指数( 式中,H′为多样性指数;Pi为第i种的个体比例);以Pielou(1949) 的公式J=H′/lnS计算小兽及其寄生蚤群落的均匀度指数(式中,J为均匀度,S为种类数);Simpson(1949) 的公式C′=∑(ni/N)2计算各群落的生态优势度(式中C′为生态优势度,ni为每一种的个体数,N为总个体数);Jaccard相似性系数公式(CJ)=j/(a+b-j),式中,j为两种景观系统共有种数,a和b分别为景观A和景观B的物种数。
根据小兽及其寄生蚤群落中各物种的捕获比例,按照徐正会等[8]提出的依据物种个体数量占样本个体总数的比例,将构成比>10%的种类定为优势种,1%~10%为常见种,<1%为稀有种。
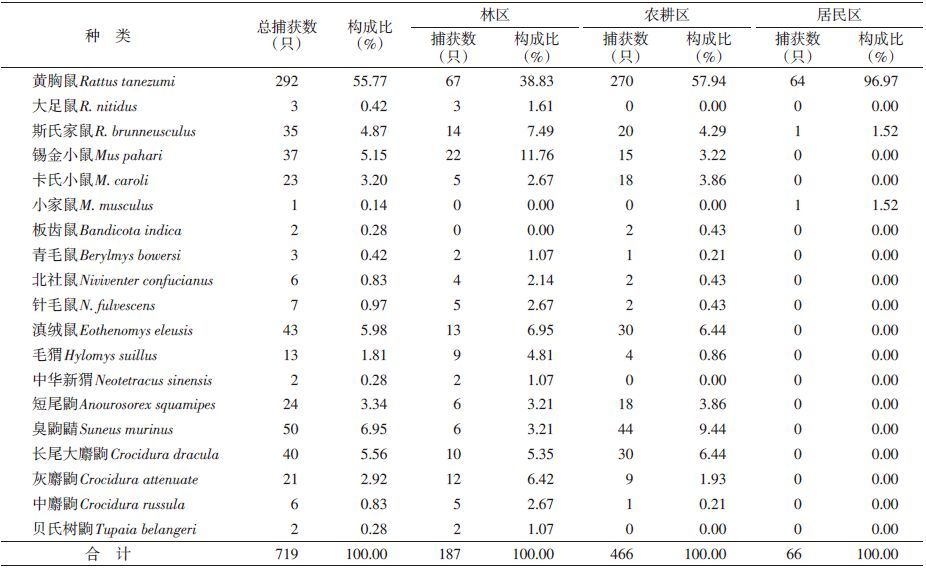
2 结 果 2.1 不同景观小兽组成本次调查共捕获小兽3目6科12属19种。其中林区3目6科11属17种,农耕区2目5科10属15种,居民区1目2科3属3种,见表 1。其中滇西南地区鼠类的优势种为黄胸鼠(Rattus tanezumi,55.77%),在3种景观内均占较大优势,说明黄胸鼠对不同景观的适应力较强,尤其在居民区为绝对优势种;常见种为臭鼩鼱(Suncus murinus,6.95%)、滇绒鼠(Eothenomys eleusis,5.98%)和长尾大麝鼩(Crocidura dracula,5.56%)等。不同景观小兽的组成、分布和小兽的捕获率不同,其物种丰富度在4~18种之间,捕获率在2.59%~7.04%之间,见表 1、2。
 |
 |
本次调查检获小兽寄生蚤共229匹,隶属5科11属14种。其中林区捕获4科8属9种,共49匹;农耕区捕获5科9属11种,共65匹;居民区为1科2属2种,共115匹。滇西南地区小兽体外寄生蚤的优势种为印属客蚤(Xenopsylla cheopis)和泸水栉眼蚤(Ctenophtha lmus lushuiensis),常见种为方叶栉眼蚤(C. quadratus)等。林区优势种为泸水栉眼蚤和方叶栉眼蚤等,常见种为无孔微棒蚤(Stiralius aporus)、滇西韧棒蚤(Lentistivalius occidentayunnanus)和偏远古蚤(Paloeopsylla remota);农耕区优势种为泸水栉眼蚤和印鼠客蚤,常见种为内曲古蚤(P. incurve)、近端远棒蚤(Aviostivalius klossi)和方叶栉眼蚤等;居民区优势种为印鼠客蚤,见表 3。
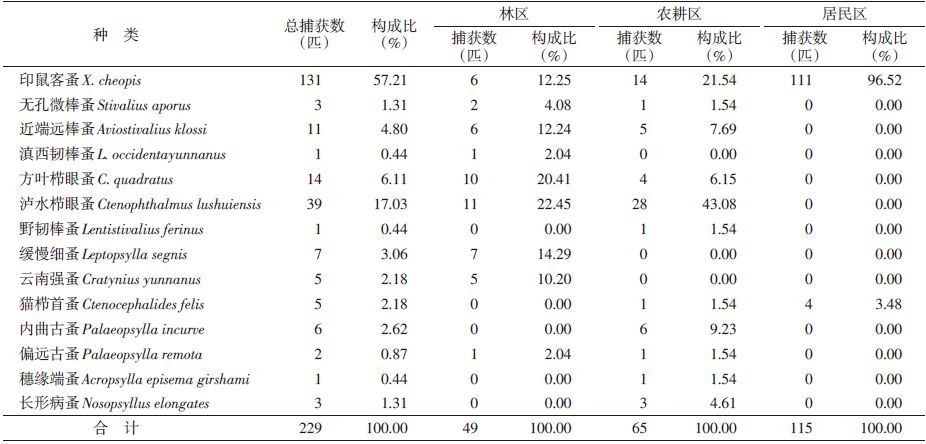 |
调查结果显示,捕获的19种小型兽类中,染有蚤类的小型兽类有8种。其中,黄胸鼠染带蚤种最多,共8种;其次是滇绒鼠和长尾大麝鼩,共3种;锡金小鼠、短尾鼩和灰麝鼩染带蚤类较单一,仅1种。捕获绝对数量以印鼠客蚤最多,主要寄生于家鼠属的鼠类,是黄胸鼠的优势种;其次是泸水栉眼蚤和方叶栉眼蚤,是滇绒鼠的优势种;其他蚤类可能是对应宿主动物的常见种或偶然染带。
本次调查小型兽类的染蚤率为12.21%,其中滇绒鼠染蚤率最高,为39.53%,其次是长尾大麝鼩和黄胸鼠,染蚤率分别为15.00%和11.47%,锡金小鼠、毛猬、短尾鼩和灰麝鼩染蚤率较低;总蚤指数为0.37,蚤指数最高的是滇绒鼠,为1.21;最低的为锡金小鼠,为0.03,其他小兽蚤指数在0.04~0.46之间,见表 4。
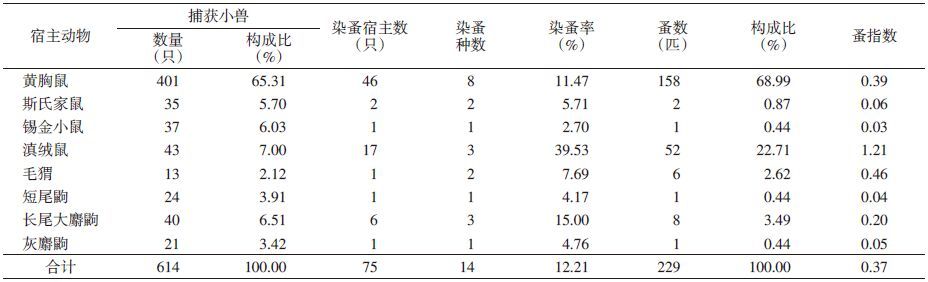 |
小兽物种多样性指数、物种丰富度以及均匀度指数均以林区最高,其次是农耕区,居民区最低。生态优势度则相反,居民区>农耕区>林区。小兽寄生蚤物种多样性指数、均匀度指数在不同景观中的分布与小兽一致,见表 5。
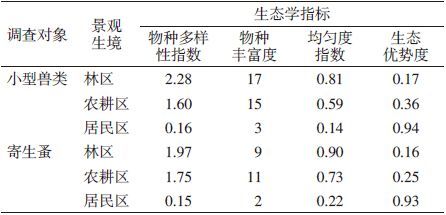 |
3种景观小型兽类群落间相似性系数有区别,其中林区与农耕区相似性系数最高为0.78,该区域农耕地大多属于山地耕地,多与林区呈交错或斑块分布,使得林区和农耕地景观小型兽类的分布相似,而居民区环境较为单一,小型兽类多与家鼠属鼠类为主,与另外两种景观中的小兽群落极不相似。小兽寄生蚤群落在不同景观中的相似性系数均不高,林区与农耕区之间蚤类群落仅为中等不相似,而居民区与其他两类景观中的蚤类群落为极不相似,说明环境对小型兽类及其寄生蚤的群落分布有一定作用,见表 6。
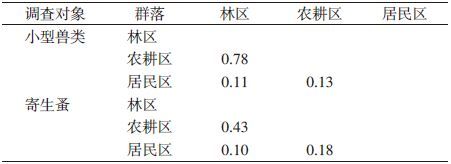 |
鼠疫自然疫源地可以看作是依附于陆生生态系统中特殊的鼠疫生物地理群落,由生态地理景观、宿主、媒介和病原体四要素组成,由于地域环境和鼠疫生态地理景观不同,导致鼠疫生物群落种群、数量、生态和遗传生物学规律有所差异[9]。从调查结果可知,不同景观小型兽类的目、科、属、种有较大区别,物种丰富度以林区居多、农耕区次之、居民区最少的分布趋势,该疫源地鼠疫主要宿主动物黄胸鼠在3种景观中均为优势种,说明其作为主要宿主对不同景观具有较强适应能力,但在不同景观中的构成有明显区别,以居民区最高;而不同景观捕获寄生蚤的组成有明显差异,林区和农耕区优势种以栉眼蚤属为主,而环境单一的居民区优势蚤种为印鼠客蚤,这也是滇西家鼠疫源地鼠疫流行多为居民区的主要原因,显示出不同景观在宿主动物和媒介蚤类的群落结构中起支配作用,鼠疫动物病及人间鼠疫流行与主要宿主(黄胸鼠)和传播媒介(印鼠客蚤)在不同景观的空间分布特征有密切关系。
滇西南家鼠鼠疫疫源地从1990年临沧市耿马县暴发流行至2007年,以1997-1999年流行最为严重,动物疫情每年均有发生,累计发生82县次,流行348点次,证实鼠疫患者130例,占全省所有病例的25.64%[10],其长期存在和发生机制,是人群、病原、宿主和媒介位于不同地域、环境梯度中或群落内的不同物种间相互关系的群落生态学问题。本次调查结果显示,在滇西南家鼠鼠疫疫源地3类景观中,小型兽类及其寄生蚤类群落多样性指数和均匀度指数呈现出从高到低依次为林区>农耕地>居民区,而优势度指数则相反的空间分布格局。根据Ostfeld和Keesing[11]研究表明,对于某些特定的传染病,媒介生物群落高多样性会降低人、畜共患病风险,滇西南林区景观中由于森林植被保护较好,环境异质性较高,小型兽类及其寄生蚤类物种多样性和均匀度指数较高,鼠疫流行的风险降低,有利于维持家鼠疫源地的自然疫源性;而居民区景观中小兽及其寄生蚤群落物种多样性和均匀度指数较低,主要宿主(黄胸鼠)和传播媒介(印鼠客蚤)优势明显,不仅是动物鼠疫流行的重要环境,也是家鼠鼠疫病例主要出现于居民区的重要原因[12, 13, 14];而农耕区各项生物学指标正好介于林区和居民区两种景观之间,说明农耕区这种景观作为鼠疫主要宿主及传播媒介生存和交流于两类景观的过渡区域,可能是鼠疫传播和流行的重要途径,在鼠疫疫源地的长期保存、流行和扩散机制中起着关键作用。
群落的相似性分析是从不同角度反映群落结构的异同,并能间接反映物种多样性的分布规律。通过对比不同景观小兽及其寄生蚤群落结构的差异,发现林区和农耕区的小型兽类及其寄生蚤群落相似程度高,与滇西南农耕地多为山坡耕地,集约化程度相对较低[15],且与林区相间交错分布有关;而居民区则与它们有明显区别,居民区作为一个特殊的动物栖息生境,受社会经济发展和人为活动的影响较大,小兽及其寄生蚤群落较为简单,鼠疫主要宿主和传播媒介的优势地位突出,既是云南省家鼠型鼠疫暴发流行的主要景观,也是鼠疫防治工作的重点区域。
影响家鼠鼠疫疫源地分布、发生和流行的因素很多,如不同地理景观和气候环境条件下,小型兽类及其寄生蚤类群落、物种多样性、尤其是重要宿主和媒介的地理分布特征、物种多度时空分布格局,以及它们在不同家鼠鼠疫疫源地的选择适应和变化趋势对鼠疫发生频率、分布规律和长期维持疫源性的综合影响等,都值得进一步探讨和研究。
| [1] | 郑宇婷,王剑,曾旭灿,等. 云南省普洱市南部山区小型兽类组成及其分布特征[J]. 中国媒介生物学及控制杂志,2014,25(6):496-501. |
| [2] | 龚正达,吴厚永,段兴德,等. 云南临沧地区蚤类群落生态学与区系研究[J]. 寄生虫与医学昆虫学报,2000,7(3):160-169. |
| [3] | 赵春花,李保华,周虹羽,等. 2003-2012年云南省景洪市鼠疫监测结果分析[J]. 疾病监测,2014,29(10):810-813. |
| [4] | 张胜勇,郭宪国,龚正达,等. 云南省21县市小兽体表蚤类群落生态及种多度分布[J]. 中国人兽共患病学报,2008,24(6):518-521. |
| [5] | 刘霖,刘正祥,杜春红,等. 2007-2012年云南省居民区鼠疫宿主动物调查及其地理分布[J]. 中华地方病学杂志,2014,33(5):517-521. |
| [6] | 詹银珠,郭宪国,左小华,等. 云南省部分地区地里纤恙螨分布调查[J]. 中华流行病学杂志,2011,32(1):13-16. |
| [7] | 钱迎倩,马克平. 生物多样性研究的原理与方法[M]. 北京:中国科学技术出版社,1994:141-167. |
| [8] | 徐正会,杨比伦,胡刚. 西双版纳片断山地雨林蚁科昆虫群落研究[J]. 动物学研究,1999,20(4):288-293. |
| [9] | 方喜业,刘起勇,许磊,等. 中国鼠疫自然疫源地分型研究Ⅷ.鼠疫自然疫源地分型生物学研究基础理念[J]. 中华流行病学杂志,2013,34(1):91-97. |
| [10] | 杨春光,赵文红,董兴齐. 云南省家鼠鼠疫流行特征与强度分析[J]. 中国媒介生物学及控制杂志,2007,18(3):226-229. |
| [11] | Ostfeld RS, Keesing F. The function of biodiversity in the ecology of vector-borne zoonotic diseases[J]. Can J Zool, 2000, 78(12): 2061-2078. |
| [12] | Guo XG, Gong ZD, Qian TJ, et al. Flea fauna investigation in some foci of human plague in Yunnan, China[J]. Acta Zootaxon Sin,2000,25(3):291-297. |
| [13] | 吴爱国,宋志忠,李天元,等. 滇西4市县居民区小哺乳动物的群落结构[J]. 兽类学报,2003,23(4): 332-338. |
| [14] | 吴爱国,李天元,冯建孟,等. 中国滇西家鼠鼠疫宿主及其媒介群落结构差异的流行病学意义[J]. 中华流行病学杂志,2008,29(4):346-350. |
| [15] | 彭尔瑞,王穗,郝莉莎,等. 云南耕地与基本农田保护现状分析及对策研究[J]. 资源与产业,2009,11(4):59-63. |