 2016, Vol. 27
2016, Vol. 27扩展功能
文章信息
- 栗冬梅
- LI Dong-mei
- 一群古老又新鲜的致病菌:巴尔通体
- A group of emerging and reemerging pathogens: Bartonella spp.
- 中国媒介生物学及控制杂志, 2016, 27(1): 84-88
- Chin J Vector Biol & Control, 2016, 27(1): 84-88
- 10.11853/j.issn.1003.4692.2016.01.027
-
文章历史
- 收稿日期: 2015-09-29



1885年8月的一天,年轻的秘鲁医生——丹尼尔·卡瑞恩(Daniel Alcides Carrión)在自己的手臂上接种了秘鲁疣(Verruga peruana)患者皮损内的液体[1],21 d后,他出现发热、寒战和骨痛等症状,在昏迷前他记录了全部发病过程,35 d后,他死于急性溶血性贫血——奥罗亚热(Oroya fever)。虽然不清楚病因是什么,但在死前卡瑞恩确信是同样的致病因子引起的秘鲁疣和奥罗亚热。后来人们以他的名字将这两种表现不同的疾病命名为卡瑞恩病(Carrion’s disease)。1905年,秘鲁的微生物学家艾尔伯特·巴顿(Alberto Barton)在患者的红细胞中发现了这种疾病的致病因子。1919年,泰勒马克·巴蒂斯梯尼(Telémaco S. Battistini)和野口英世(Hideyo Noguchi)分离出病原体,为了纪念巴顿首次发现这种微生物,将其命名为杆菌样巴尔通体(Bartonella bacilliformis)[2, 3],这就是巴尔通体属的第一个种。在随后的几十年中,虽然陆续发现了五日热巴尔通体、汉赛巴尔通体和伊丽莎白巴尔通体等,但均归类为罗莎利马体属。直到20世纪90年代,随着分子分类学方法在细菌分类鉴定中的应用,巴尔通体的分类学位置重新调整[4],去除罗莎利马体和格拉汉姆体属,归为巴尔通体属,移出立克次体目、归入根瘤菌目,理顺了巴尔通体科、属的分类关系。此后,陆续不断地有新种巴尔通体被发现命名,这些巴尔通体宿主动物广泛分布,包括猫、犬、鼠、牛和蝙蝠[5],甚至海洋哺乳动物等[6]。自此,巴尔通体研究进入了一个新时代,巴尔通体也被看做是新发传染病病原菌受到关注,其生物学、流行病学、致病机制和系统发育分类等研究快速发展,全基因组测序研究的不断推进,更使得巴尔通体研究进入了后基因组时代。为此,我们利用GoPubMed数据库,对巴尔通体相关文献进行计量分析,梳理巴尔通体研究脉络,概述研究进展,为进一步了解和深入研究提供参考信息。
1 文献搜索概述在PubMed检索系统中搜索巴尔通体文献,起止时间为1926年5月至2015年3月。为使结果更清晰、直观,采用与PubMed有着共同数据信息但同时又具备多种统计分析功能的GoPubMed作为检索工具。为保证检索结果全面、准确,输入检索词“Bartonella”,得到文献3 043篇,综述349篇;输入检索词“Rochalimaea”(因巴尔通体在1992年前曾被命名为罗莎利马体),得到文献3 135篇,包括上述检索词“Bartonella”所得全部文献,综述364篇,多出92篇文献主要为1994年以前描述Rochalimaea内容的文献。其中,鼠传巴尔通体是近年来研究较多、发展较快的领域,自1964年至今有文献290篇(检索词为Bartonella和rodent),1993年后研究报道迅速上升,新鉴定的鼠传巴尔通体种类也在快速增加,文中将详细论述。
2 全球文献的时间分布在PubMed中输入检索词“Bartonella”,截止到2015年3月共计检索出相关文献3 043篇,其中综述349篇,最早文献出现在1926年[2],1992年以前每年文献量在20篇以下,此后呈迅速上升趋势,至2012年最多,收录171篇。GoPubMed统计的巴尔通体相对研究兴趣指数(relative research interest,RRI)逐年提高,从1994年以后增长幅度较大,至2012年达到最高水平,近几年有下降趋势(图 1)。最早期的文献是关于杆菌样巴尔通体(B. bacilliformis)相关内容,有222篇文献;汉赛巴尔通体相关文献有1 590篇;五日热巴尔通体相关文献有478篇;这3种巴尔通体研究文献占全部文献的75.25%。

|
| 图 1 巴尔通体相关文献年代分布 Figure 1 Temporal distribution of Bartonellae related literatures |
全球除南极洲外各大洲均有巴尔通体研究报道,分布于86个国家,以欧洲和北美洲研究最多(图 2)。文献量最多的前10位为美国(898篇)、法国(335篇)、德国(137篇)、日本(97篇)、意大利(87篇)、英国(83篇)、西班牙(73篇)、瑞士(63篇)、巴西(52篇)和以色列(43篇)。中国发表文献38篇,位居第11位,自2004年起呈增多趋势,晚于国际上近10年。实际上,国内相关研究报道起始于20世纪70年代,主要是猫抓病相关的病例报道[7, 8],因为中文文献尚未被收录入PubMed数据库,故未被统计在内。

|
| 图 2 巴尔通体相关文献的全球分布(引自GoPubMed) Figure 2 Geographical distribution of Bartonellae related literatures (quoted from GoPubMed) |
巴尔通体相关文献投稿2篇以上论文的期刊共计371种,发表相关文献最多的期刊是Journal of Clinical Microbiology,其他还有Emerging Infectious Diseases、Vector-borne and Zoonotic Diseases和Clinical Infectious Diseases等,排名前10位的期刊文献量占文献总量(3 043篇)的20.8%,见表 1。在高影响因子期刊中,发表巴尔通体相关论文最多的是Journal of Experimental Medicine(37篇)。国际权威杂志如Nature、Science、Lancet和New England Journal of Medicine等发表相关研究分别为6篇、3篇、23篇和11篇。中国相关论文发表较高影响因子的英文期刊有Emerging Infectious Diseases(3篇)、Journal of Bacteriology(1篇)、PLoS Negl Trop Dis(1篇)和Bioinformatics(1篇)等。
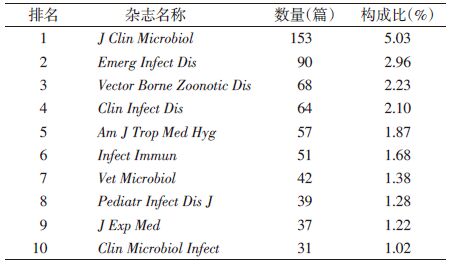 |
依据GoPubMed给出的作者情况,法国马赛医学院的Raoult发表巴尔通体相关文章174篇,排名第一;其次是美国、瑞士和英国等的研究者有较高的文献量,>20篇。从图 3中的连线和其线条的粗细,可知研究者们之间的合作关系和密切程度。多产作者们形成两大主要合作网络,一个是以法国Raoult为中心的欧洲合作网络,另一个是Chomel(63篇)和Breitschwerdt(70篇)等的美国合作网络,他们研究内容以病原分离及分子流行病学调查为主。近年瑞士科学家Dehio(35篇)的团队在巴尔通体Ⅳ型分泌系统的分子结构、组成和功能等方面做了大量研究[9, 10, 11],更是从基因组进化的角度发现了其与宿主适应及特异性密切相关[12],填补了巴尔通体分子致病机制的空白。此外,在继国外研究发现食蟹猴可携带五日热巴尔通体后[13],中国学者相继在猕猴中分离出五日热巴尔通体,在其新宿主发现方面做了贡献,突破了以往认为人类是唯一宿主的认识,为五日热巴尔通体动物来源提供了证据[14, 15, 16]。

|
| 注:1~6表示合作者数量。 图 3 研究人员网络关系(引自GoPubMed) Figure 3 Network of senior authors (quoted from GoPubMed) |
第9版《伯杰氏系统细菌学手册》(2004年)发布的巴尔通体有21种[4],此后不断有新种被发现和命名。到目前为止,新增12种,主要是一些鼠传巴尔通体,其中罗莎利马巴尔通体可引起人类发热性疾病(表 2)。在此,基于gltA(326 bp)、rpoB(771 bp)和ftsZ(766 bp)基因多位点串联序列、应用TOPALi 2.5软件(Biomathematics and Statistics Scotland,Edinburgh,UK)中贝叶斯分析MCMC算法(HKY+I+G模型,8 000 000 generations,25% burn-in)构建巴尔通体属的系统发育树,除白足鼠巴尔通体(B. peromysci)和鼹鼠巴尔通体(B. talpae)外,其余29种巴尔通体均归在相应的基因群(图 4)。该系统发育树将巴尔通体分为6个群,分别以不同颜色标注。其中,鼠类宿主和反刍动物宿主巴尔通体有明显的聚集,还有一些种无明确位置。使用不同基因、不同方法所构建的巴尔通体属系统发育树拓扑结构有一定的差异,使某些种在不同的系统发育树中的位置有差异[9, 21, 23],目前这些树均是基于几个基因的信息量,是否能够代表物种的进化关系有待商榷,或许全基因组系统发育树的构建能够给我们带来更多信息和解析。


|
| 图 4 基于gltA、rpoB和ftsZ基因串联序列的巴尔通体属系统发育树 Figure 4 Bayesian phylogeny of the genus Bartonella based on the concentration of gltA, ftsZ and rpoB |
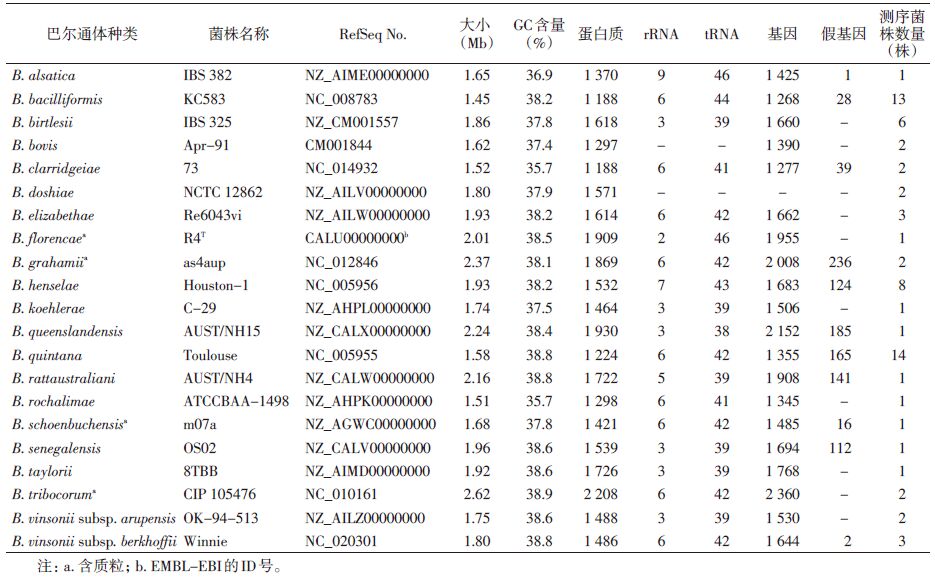
目前,有21种巴尔通体基因组已被测序,有些已可以获取全基因组序列(表 3)。与大多数细菌(如Escherichia coli>4 Mb)相比,巴尔通体有1个较小的环状基因组,大小范围是1.45 Mb(1 268个编码基因)~2.62 Mb(2 360个编码基因),平均为1.85 Mb,与立克次体和无形体(约1.1 Mb~1.2 Mb)相似。巴尔通体核心基因组有959个基因[16],全部编码部分估计不低于70%,前导链有大量的G和T,显示出很强的GC偏态[24],GC含量为35.70%~38.90%,其中4种巴尔通体携带质粒。在大多数基因组中存在前噬菌体和基因组岛,相比人类特异性病原五日热巴尔通体和杆菌样巴尔通体则不存在这类结构。在巴尔通体基因组中存在大量重排序列和重组事件,使得基因组呈现出扩张态势,并通过获取T4SS系统的策略适应各种动物宿主[12]。为平衡基因组大小,衰减也是巴尔通体特征之一,专性寄生(如五日热巴尔通体和杆菌样巴尔通体)导致其较窄的生态位,促进了基因丢失这一过程[25],这一特征与专性人类寄生密切相关,在伤寒沙门菌(Salmonella typhi)、普氏立克次体(Rickettsia prowazekii)和麻风杆菌(Mycobacterium leprae)也发现这种进化现象。在磷脂代谢通路中,相关基因(glpK、the Glp system和gpsA)的丢失和重新获得存在于巴尔通体中,再次揭示水平转移(HGT)在巴尔通体生态适应性进化中起重要作用[26]。
有人认为杆菌样巴尔通体引起的疾病可以追溯到美洲的前印加时代[27],虽然这种疾病在未治疗的患者中致死率高达44%~88%[28],但是仅流行于南美洲安第斯山脉地区,使得巴尔通体一直以来并未引起广泛关注。直到20世纪80年代,分子生物学方法应用于巴尔通体分类鉴定,使得人们重新认识了这类病原菌与相关疾病之间的关系。在新方法应用后,原本不认识的微生物甚至是致病微生物大量出现,使我们对疾病有了新的认识。原本认为无致病性的物种也出现了病例报道,提示病原菌的概念是相对的,关键看主体是谁。对于病原菌,寄主是他们的生态栖息地,而对于寄主,如果产生危害,就可以认为是病原菌。但如果进化为相互适应,如巴尔通体寄生于啮齿动物中,通过减少基因组而降低对宿主的毒性,实施妥协策略,以更利于生存繁殖,即可相安无事。当生态条件发生变化时,则可能具有致病性,如变换宿主类型时,对新宿主可能产生危害,即由原来的非致病性转变为致病性。很多鼠传巴尔通体即是这类微生物,由于人类偶然接触感染而致病,并且无特征性临床表现,难于诊断。这种情况,随着人类与自然接触愈发密切而会越来越多,面临的传染病流行情况会更加复杂,防控任务更加艰巨。
志谢 原上海医科大学温廷桓教授对本文中巴尔通体中文译名翻译给予指导与帮助,特此志谢
| [1] | Schultz MG. Who is this man?[J].Emerg Infect Dis,2010,16(6):1025-1027. |
| [2] | Noguchi H, Battistini TS. Etiology of oroya feverⅠ. cultivation of Bartonella bacilliformis[J].J Exp Med,1926,43(6):851-864. |
| [3] | Noguchi H. The etiology of verruga peruana[J].J Exp Med, 1927,45(1):175-189. |
| [4] | 文心田,于恩庶,徐建国,等. 当代世界人兽共患病学[M].成都:四川科学技术出版社,2011:952-966. |
| [5] | Kamani J, Baneth G, Mitchell M, et al. Bartonella species in bats (Chiroptera) and bat flies (Nycteribiidae) from Nigeria,West Africa[J].Vector Borne Zoonotic Dis,2014,14(9):625-632. |
| [6] | Harms CA, Maggi RG, Breitschwerdt EB, et al. Bartonella species detection in captive, stranded and free-ranging cetaceans[J].Vet Sci,2008,39(6):59. |
| [7] | 栗冬梅,张建中,刘起勇. 中国巴尔通体与相关疾病的研究进展[J].中国人兽共患病学报,2008,24(8):762-765. |
| [8] | 黄儒婷,刘起勇. 中国内地猫抓病流行病学研究分析[J].中华预防医学杂志,2009,43(5):442-444. |
| [9] | Engel P, Salzburger W, Liesch M, et al. Parallel evolution of a typeⅣ secretion system in radiating lineages of the host-restricted bacterial pathogen Bartonella [J].PLoS Genet,2011,7(2):e1001296. |
| [10] | Seubert A, Hiestand R, de la Cruz F, et al. A bacterial conjugation machinery recruited for pathogenesis[J].Mol Microbiol,2003,49(5):1253-1266. |
| [11] | Schmid MC, Schulein R, Dehio M, et al. The VirB type Ⅳ secretion system of Bartonella henselae mediates invasion, proinflammatory activation and antiapoptotic protection of endothelial cells[J].Mol Microbiol,2004,52(1):81-92. |
| [12] | Engel P, Dehio C. Genomics of host-restricted pathogens of the genus Bartonella[J].Genome Dyn,2009,6:158-169. |
| [13] | O'Rourke LG,Pitulle C,Hegarty BC, et al. Bartonella quintana in cynomolgus monkey (Macacafascicularis)[J].Emerg Infect Dis,2005,11(12):1931-1934. |
| [14] | Huang R,Liu QY,Li GP,et al. Bartonella quintana infections in captive monkeys,China[J].Emerg Infect Dis,2011,17(9):1707-1709. |
| [15] | Li H, Bai JY, Wang LY, et al. Genetic diversity of Bartonella quintana in macaques suggests zoonotic origin of trench fever[J].Mol Mcol,2013,22(8):2118-2127. |
| [16] | Li H, Liu W, Zhang GZ, et al. Transmission and maintenance cycle of Bartonella quintana among rhesus macaques, China[J].Emerg Infect Dis,2013,19(2):297-300. |
| [17] | Gundi VAKB, Taylor CT, Raoult D, et al. Bartonella rattaustraliani sp. nov.,Bartonella queenslandensis sp. nov. and Bartonella coopersplainsensis sp. nov., identified in Australian rats[J].Int J Syst Evol Microbiol,2009,59(12):2956-2961. |
| [18] | Inoue K, Kabeya H, Shiratori H, et al. Bartonella japonica sp. nov. and Bartonella silvatica sp. nov., isolated from Apodemus mice[J].Int J Syst Evol Microbiol,2010,60(4):759-763. |
| [19] | Eremeeva ME, Gerns HL, Lydy SL, et al. Bacteremia, fever, and splenomegaly caused by a newly recognized Bartonella species[J].N Engl J Med,2007,356(23):2381-2387. |
| [20] | Sato S, Kabeya H, Fujinaga Y, et al. Bartonella jaculi sp. nov., Bartonella callosciuri sp. nov., Bartonella pachyuromydis sp. nov. and Bartonella acomydis sp. nov., isolated from wild rodentia[J].Int J Syst Evol Microbiol,2013,63(5):1734-1740. |
| [21] | Mediannikov O, El Karkouri K, Robert C,et al. Non-contiguous finished genome sequence and description of Bartonella florenciae sp. nov.[J].Stand Genomic Sci,2013,9(1):185-196. |
| [22] | Mediannikov O, El Karkouri K, Diatta G, et al. Non-contiguous finished genome sequence and description of Bartonella senegalensis sp. nov.[J].Stand Genomic Sci,2013,8(2):279-289. |
| [23] | Saenz HL, Engel P, Stoeckli MC, et al. Genomic analysis of Bartonella identifies typeⅣ secretion systems as host adaptability factors[J].Nat Genet,2007,39:1469-1476. |
| [24] | Alsmark CM, Frank AC, Karlberg EO, et al. The louse-borne human pathogen Bartonella quintana is a genomic derivative of the zoonotic agent Bartonella henselae[J].Proc Natl Acad Sci USA,2004,101(26):9716-9721. |
| [25] | Sällström B, Andersson SG. Genome reduction in the α-Proteobacteria[J].Curr Opin Microbiol,2005,8(5):579-585. |
| [26] | Zhu QY, Kosoy M, Olival KJ, et al. Horizontal transfers and gene losses in the phospholipid pathway of Bartonella reveal clues about early ecological niches[J].Genome Biol Evol, 2014,6(8):2156-2169. |
| [27] | Schultz MG. A history of Bartonellosis (Carrión's disease)[J].Am J Trop Med Hyg,1968,17(4):503-515. |
| [28] | Maguiña C, Guerra H, Ventosilla P. Bartonellosis[J].Clin Dermatol,2009,27(3):271-280. |