 2015, Vol. 26
2015, Vol. 26扩展功能
文章信息
- 靳铁治, 吴瑞, 陈海龙, 陈晓宁, 马超锋, 王开锋
- JIN Tie-zhi, WU Rui, CHEN Hai-long, CHEN Xiao-ning, MA Chao-feng, WANG Kai-feng
- 西安市肾综合征出血热宿主动物调查及病原分析
- Investigation and analysis on host animals and pathogen of hemorrhagic fever with renal syndrome in Xi'an city, Shaanxi, China
- 中国媒介生物学及控制杂志, 2015, 26(4): 379-382
- Chin J Vector Biol & Control, 2015, 26(4): 379-382
- 10.11853/j.issn.1003.4692.2015.04.013
-
文章历史
- 收稿日期:2015-02-27




2 西北大学生命科学学院, 陕西西安710069;
3 西安市疾病预防控制中心
2 The College of Life Sciences, Northwest University, Xi'an 710069, Shaanxi Province, China;
3 Xi'an Center for Disease Control and Prevention
肾综合征出血热(HFRS)是一种主要由鼠类传播的自然疫源性传染病[1],其病原汉坦病毒属于布尼亚病毒科的汉坦病毒属,中国的HFRS病例主要由黑线姬鼠(Apodemus agrarius)携带的汉滩型病毒(Hantaan virus,HTNV)和由褐家鼠(Rattus norvegicus)携带的汉城型病毒(Seoul virus,SEOV)引起[2]。2007年以来,陕西省HFRS疫情逐年上升,至2010年发病高达2356例,死亡28例,位居全国各省发病数前列。西安市HFRS 发病占陕西省的90%以上[3]。近年来,西安市HFRS发病例数持续上升,老疫区逐渐扩大,新的疫区不断出现,汉坦病毒不断出现新的血清型,并不断在新的宿主动物体内被发现,传播风险增大[4, 5, 6]。鼠类作为HFRS的主要宿主动物,其种类分布及变化对HFRS的流行起到关键作用[7]。近来,不断有人研究宿主动物生态特征与传播疾病的关系,Clay等[8]认为宿主动物生物多样性在辛诺柏病毒传播过程中有重要作用。Kathleen等[9]认为生物多样性丰富可降低莱姆病的传播风险。Hu等[10]认为九华山林区由于生境汉坦病毒的传播率比较低,可能是由于林区生物多样性更加丰富。随着西安市城镇化进程加快,生境破碎化逐渐严重,鼠形动物种类和栖息地随之发生变化。因此,为进一步了解西安市HFRS宿主动物的种类分布及变化,了解当前鼠形动物携带的汉坦病毒型别,为HFRS的防控政策提供更加科学、有效的依据,本研究对西安市的鼠形动物进行了HFRS宿主动物调查。 1 材料与方法 1.1 实验动物
根据2011年西安市HFRS的发病情况,并根据鼠形动物的活动特征,于2012年1-12月对西安市的HFRS 宿主动物进行了调查,选择2011年度发病率较高的3个区(长安区、临潼区和户县)作为研究区域。 1.2 研究方法
1.2.1 监测方法
采用夹夜法,将研究区域划分为居民区和野外2种生境,又将居民区生境分为一般场所(家属院)和特殊场所(菜、果品市场),野外生境分为农田、撂荒地和林地。野外夹距5 m×5 m,居民区每15 m2布放1夹,用生花生拌香油作诱饵,傍晚布夹,次日清晨收回,统计捕获率。对鼠类进行鼠种鉴定和常规指标的测量[11, 12],同时计算鼠种构成,并在无菌条件下剖取鼠肺,放入带有编号的冷冻塑料管内,置于液氮罐以备病毒检测。 1.2.2 病毒检测
利用组织研磨器(Tissue lyserLT,Qiagen Hildon,Germany)均质动物肺脏后用Trizol从肺组织中提取RNA,操作过程按照Trizol试剂(美国Invitrogen公司)使用说明书进行。提取的RNA空气干燥后溶于DEPC(焦碳酸二乙酯)处理水中,利用六碱基随机引物合成cDNA(Fermentas,Canada),按照《全国肾综合征出血热监测方案(试行)》中的汉坦病毒基因分型方法进行检测。所用引物见表 1。
1.3 数据处理
采用SPSS 19.0统计软件进行数据分析,所有数据首先通过One-sample K-S检验数据正态性,采用t 检验分析不同月份间鼠密度及鼠类带病毒率是否存在差异;一般线性模型分析不同生境鼠形密度的差异,将显著水平设为P<0.05。 2 结果 2.1 宿主动物种类构成
本次调查捕获的鼠形动物有2目4科6属12种480只,有多齿鼩鼹(Uropsilussoricipes)、北小麝鼩(Crocidura suaveolens)、灰麝鼩(C. attenuata)、北社鼠(Niviventer confucianus)、黄胸鼠(Rattus tanezumi)、褐家鼠、小家鼠(Mus musculus)、高山姬鼠(Apodemus chevrieri)、大林姬鼠(A. peninsulae)、中华姬鼠(A. draco)、黑线姬鼠和大仓鼠(Tscharskiatriton)。
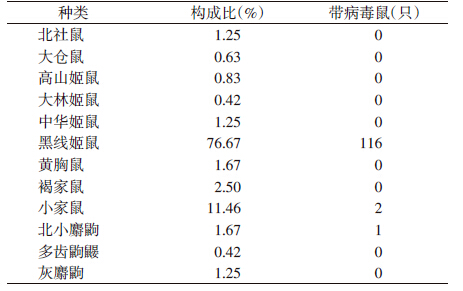
在捕获的480只鼠形动物中,黑线姬鼠为优势种,占捕获总数的76.67%;携带汉坦病毒的种类有3种,分别为黑线姬鼠、小家鼠和北小麝鼩(表 2)。
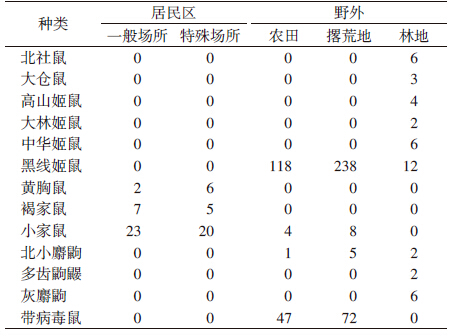
不同生境调查中,居民区以小家鼠为优势种;野外黑线姬鼠在农田、撂荒地、林地3种小生境中均为优势种(表 3)。
 |
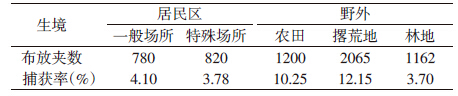
本次调查共布设鼠夹累计6027夹夜,捕获鼠形动物480只。不同生境动物密度差异有统计学意义(F=4.555,P<0.05);野外鼠形动物密度显著高于居民区(Z=6.94,P<0.01),其中居民区布设1600夹,捕获鼠形动物63只,捕获率为3.94%;野外布设4427夹,捕获鼠形动物417只,捕获率为9.42%。在野外生境中,不同小生境密度差异有统计学意义(F=7.892,P<0.05),撂荒地的密度最高,为12.15%,其次为农田和林地,分别为10.25%和3.70%(表 4)。主要宿主动物黑线姬鼠密度从5月急剧上升,8月达到最高,11月后明显下降(图 1)。

|
| 图 1 不同月份鼠形动物密度及带病毒情况 Figure 1 Density and virus-carrying rate of the animal in different months |
本次调查共捕获鼠形动物480只,以鼠肺为样品进行汉坦病毒检测,在119个样品中汉坦病毒的检测结果为阳性,鼠形动物的汉坦病毒阳性率为24.79%;其中,捕获黑线姬鼠368只,在116份鼠肺中检测结果为阳性,带病毒率为31.52%(表 2、3)。不同月份间鼠形动物带病毒率差异有统计学意义(t=5.894,P<0.05),鼠形动物带病毒率8月达到峰值,10月次之,11月后迅速下降,1月降至最低;不同生境动物携带汉坦病毒差异有统计学意义,在居民区未检测出阳性鼠肺,野外检测到119份阳性鼠肺,其中黑线姬鼠116份,小家鼠2 份,北小麝鼩1份,分别占带病毒鼠的97.48%、1.68%和0.84%。
对119份阳性鼠肺进行基因型别检测发现,所有样本均为Ⅰ型(汉滩型)。 3 讨论
宿主动物为疾病传播的重要环节,过去的研究表明,HFRS的宿主动物主要为鼠形动物,因此,研究区域性鼠形动物的种类构成、密度变化及动物携带病毒情况,可为区域性HFRS的预防政策提供科学依据。本研究调查结果表明,西安市野外鼠密度显著高于居民区,且带病毒鼠均捕获于野外,因此为了控制HFRS的流行,如何控制野外尤其是农田及撂荒地的鼠形动物密度尤为重要。从鼠形动物物种构成看,本次调查共统计到2目4科6属12种鼠形动物,主要为鼠科动物,鼠科动物中又以姬鼠属为主,家鼠属次之,在姬鼠属动物中,黑线姬鼠所占比例最高,约占全部鼠形动物的76.67%,为优势物种,其次为小家鼠和褐家鼠。本次调查到的12种鼠形动物,带病毒鼠全部发现于野外,野外数量上黑线姬鼠处于绝对优势,其余种类合计约占全部鼠形动物的12.00%。区域性生物多样性结构相对脆弱,群落多样性程度不高,优势度明显可能是高带病毒率的重要成因之一。黑线姬鼠作为区域性最主要带病毒鼠,其在群落结构中比例较高,种群交流将更加便利,鼠间病毒的传播机会将大大加强,增加了人群感染HFRS的概率。本次检测到的汉坦病毒阳性鼠肺中,97.48%为黑线姬鼠,仅有2个小家鼠样本和1个北小麝鼩样本检测结果为阳性。Plyusnin和Morzunov[14]认为汉坦病毒和宿主间有比较紧密的对应关系,一般一种宿主只携带一种病毒,因此宿主动物的物种类别决定其携带的汉坦病毒型别和人间HFRS流行强度。本地区在黑线姬鼠、小家鼠、北小麝鼩体内检测到的均为Ⅰ型(汉滩型),与李恒新等[15]在褐家鼠和小家鼠中也检测到Ⅰ型(汉滩型)病毒基本一致。一般认为,小家鼠携带的为Ⅱ型(汉城型),本次在其体内检测到Ⅰ型病毒(汉滩型),李恒新等[15]认为病毒核酸检测由于反转录聚合酶链反应敏感、特异,但也很容易污染产生假阳性。那么究竟是发生了病毒溢出还是由于其他原因影响所致,以及小家鼠和北小麝鼩是否已经成为区域性宿主动物等问题尚待进一步研究。谭新等[16]研究证明西安市小家鼠中检测到携带Ⅱ型(汉城型)抗原,并且从患者体内检测到汉城病毒抗体;蔡正华等[3]报道西安市在褐家鼠中检测到Ⅱ型(汉城型),也检测到过Ⅰ型病毒(汉滩型)。但上述报道中家鼠型汉城病毒所占比例均较低,另外先前部分研究是利用免疫荧光染色抗原检测,染色后对阳性的判断标准存在一定的主观性,结果可能出现假阳性。结合近年来的调查结果,我们更倾向于认为:西安市目前仍然为姬鼠型疫区。鼠形动物密度及带病毒率和HFRS的发病有关联性[17]。蔡正华等[3]认为西安市的HFRS发病呈现双峰特征,即11月的大高峰和6月的小高峰。从本次的调查结果来看,鼠形动物密度和带病毒率与人间HFRS疫情2个高峰稍有不同,鼠密度月份变化呈正态分布,密度变化表现为一个高峰,鼠形动物密度从5月显著上升,8月密度达到最高,维持较高密度到11月,从11月后密度出现显著下降,无明显的双峰特征。周旭东等[18]报道,农田鼠形动物种群数量随着季节变化而消长。调查结果表明,西安市的带病毒鼠主要分布于农田和撂荒地,当地4月气温出现显著回升,鼠形动物活动逐渐增多,6月和10月分别为当地每年的小麦和玉米收获季节,农作物收获后,鼠类食物来源趋于更加丰富,并且局部生境由于耕作被破坏,导致鼠形动物出现近距离迁移聚集,小环境密度上升,这样更有利于其种群交流与繁殖,造成种群密度上升,与张建漂等[19]和Adler[20]的研究结果相一致:鼠类的栖息环境和食物来源发生变化可以引起种群的暴发;另外,一定程度的干扰可使鼠形动物种群扩大也符合中度干扰假说(intermediate disturbance hypothesis),这可能也是8月鼠密度达到最高的原因之一,6月和10月下旬为当地收获季节,也使人群和鼠形动物接触的可能性增大。因此认为当地11月和6月出现的两次人间疫情高峰可能主要与农田劳作相关。
综上所述,随着西安市近年城市变迁范围逐渐增大,鼠形动物群落结构面临干扰加剧,这些将使西安市的HFRS流行状况更加复杂。进一步研究家鼠属动物携带病毒情况,确认病毒基因型别,长期监测害鼠的群落结构变化将有利于实时掌控HFRS的传播特征,为HFRS的防控提供有效技术支撑。适时灭鼠是HFRS防控的重要环节,当地4月相对害鼠密度较低,害鼠活动开始增多,因此,本研究建议4月对当地重点疫区进行害鼠防控工作,尤其是农田、撂荒地等生境的防控,这样可以有效降低鼠形动物密度,减少居民感染HFRS的概率。另外,疫苗接种仍然是防控HFRS的最重要途径。
志谢陕西省动物研究所张广平、常罡,西安市疾病预防控制中心蔡正华、郑海潮、李焱等,陕西师范大学博士研究生张博参与本项工作,特此志谢
| [1] | Zhang YZ,Zou Y,Fu ZF,et al. Hantavirus infections in humansand animals,China[J]. Emerg Infect Dis,2010,16(8):1195-1203. |
| [2] | Zuo SQ,Zhang PH,Jiang JF,et al. Seoul virus in patients androdents from Beijing,China[J]. Am J Trop Med Hyg,2008,78 (5):833-837. |
| [3] | 蔡正华,李倩,于立芬,等. 西安市2000-2010年肾综合征出血热发病趋势分析[J]. 中国人兽共患病学报,2012,28(7):750-753. |
| [4] | 雷新耀,杨海,颉戈鹏. 陕西省地貌特征与汉坦病毒分布关系研究[J]. 中国公共卫生管理,2009,25(5):532-534. |
| [5] | Jiang JF,Zhang WY,Yao K,et al. A new Hantaan-like virus in rodents (Apodemus peninsulae) from Northeastern China[J].VirusRes,2007,130(1/2):292-295. |
| [6] | 靳铁治,马超锋,郑海潮,等. 西安市首次发现北小麝鼩携带汉坦病毒核酸[J]. 中国病原生物学杂志,2011,6(8):565-566. |
| [7] | 郑智民,姜志宽,陈安国. 啮齿动物学[M]. 上海:上海交通大学出版社,2008:460-463. |
| [8] | Clay CA, Lehmer EM, Jeor SS, et al. Sin nombre virus and rodent species diversity: a test of the dilution and amplification hypotheses[J]. PLoS One,2009,4(7):e6467. |
| [9] | Kathleen L,Shannon TKD,Michael JN,et al. Impact of host community composition on Lyme disease risk[J]. Ecology,2008,89(10):2841-2849. |
| [10] | Hu XQ,Li SG,Liu H,et al. Diversity and distribution of host animal species of Hantavirus and risk to human health in Jiuhua Mountain area,China[J]. Biomed Environ Sci,2014,27(11):849-857. |
| [11] | 潘清华,王应祥,岩崑. 中国哺乳动物彩色图鉴[M]. 北京:中国林业出版社,2007:15-27,296-333. |
| [12] | 郑生武,李保国. 中国西北地区脊椎动物系统检索与分布[M]. 西安:西北大学出版社,1999:314-321. |
| [13] | 王世文,杭长寿,王华,等. 我国汉坦病毒基因型和基因亚型的分布研究[J]. 病毒学报,2002,18(3):211-216. |
| [14] | Plyusnin A,Morzunov SP. Virus evolution and genetic diversity of hantaviruses and their rodent hosts[J]. Curr Top Microbiol Immunol,2001,256:47-75. |
| [15] | 李恒新,马超锋,左署青,等. 西安地区2009年鼠类携带汉坦病毒的基因特征研究[J]. 西安交通大学学报:医学版,2011,32(4):433-437,448. |
| [16] | 谭新,肖丹,闫永平. 西安市户县1971-2010年肾综合征出血热疫情分析[J]. 中国媒介生物学及控制杂志,2012,23(6):577-580. |
| [17] | 靳铁治,郑海潮,李劲松,等. 黑线姬鼠种群结构与携带汉坦病毒相关性[J]. 生物学杂志,2012,29(2):22-23,50. |
| [18] | 周旭东,张永军,黄健,等. 新疆甘家湖自然保护区啮齿动物群落结构与时间动态分析[J]. 动物学杂志,2004,39(6):58-61. |
| [19] | 张建漂,杨孔,李平. 北川地震灾区农田鼠类种群动态初步调查[J]. 中国媒介生物学及控制杂志,2011,22(2):121-123. |
| [20] | Adler GH. Imapacts of resource abundance on populations of a tropical forest rodent[J]. Ecology,1998,79(1):242-254. |