 2015, Vol. 26
2015, Vol. 26扩展功能
文章信息
- 李婵, 姚苹苹, 朱函坪, 徐芳, 杨章女, 岳明, 谢荣辉, 孙一晟, 徐致遥, 王长军, 张云
- LI Chan, YAO Ping-ping, ZHU Han-ping, XU Fang, YANG Zhang-nv, YUE Ming, XIE Rong-hui, SUN Yi-sheng, XU Zhi-yao, WANG Chang-jun, ZHANG Yun
- 东南沿海地区汉坦病毒的遗传进化分析
- Phylogenetic analysis of Hantavirus in the South-east coastal area of China
- 中国媒介生物学及控制杂志, 2015, 26(3): 275-278
- Chin J Vector Biol & Control, 2015, 26(3): 275-278
- 10.11853/j.issn.1003.4692.2015.03.014
-
文章历史
- 收稿日期:2014-12-25




2. 南京医科大学第一附属医院;
3. 温州医科大学;
4. 南京军区军事医学研究所
2 The First Affliated Hospital with Nanjing Medical Univercity;
3 Wenzhou Medical University;
4 The Institute of Militalry Medicine, Nanjing Command
肾综合征出血热(HFRS)是一种疫区分布广、病死率较高的自然疫源性疾病。最初为汉滩型(HTN),之后又出现汉城型(SEO)疫区,目前HTN和SEO混合型大量存在[1]。我国东南沿海地区HFRS病例数较多,据中国疾病预防控制信息系统统计,近年浙江、江苏、福建等省发病人数有上升趋势。
汉坦病毒(Hantavirus,HV)是HFRS的病原体,属布尼亚病毒科,其基因结构分为大(L)、中(M)、小(S)3个片段,分别编码病毒的RNA聚合酶、糖蛋白(GP)G1和G2以及核衣壳蛋白(NP)[2]。本研究收集浙江、福建省和上海市汉坦病毒部分M基因序列,应用Mega 4.0和DNAStar软件包进行系统发生分析,以邻位相连法(NJ)构建系统发生树,分析东南沿海地区与其他地区汉坦病毒的基因差异。
1 材料与方法 1.1 序列收集用于比较分析的上海市、福建和浙江省的汉坦病毒部分M基因序列来自公开资料[3, 4, 5, 6, 7];另外用于本研究分析的HTN国际标准株76-118 分离自韩国的黑线姬鼠(Apodemus agrarius),M基因的GenBank编号为M14627; SEO国际标准株80-39分离自韩国的褐家鼠(Rattus norvegicus),M基因的GenBank编号为S477l6,本研究的毒株信息见表 1。
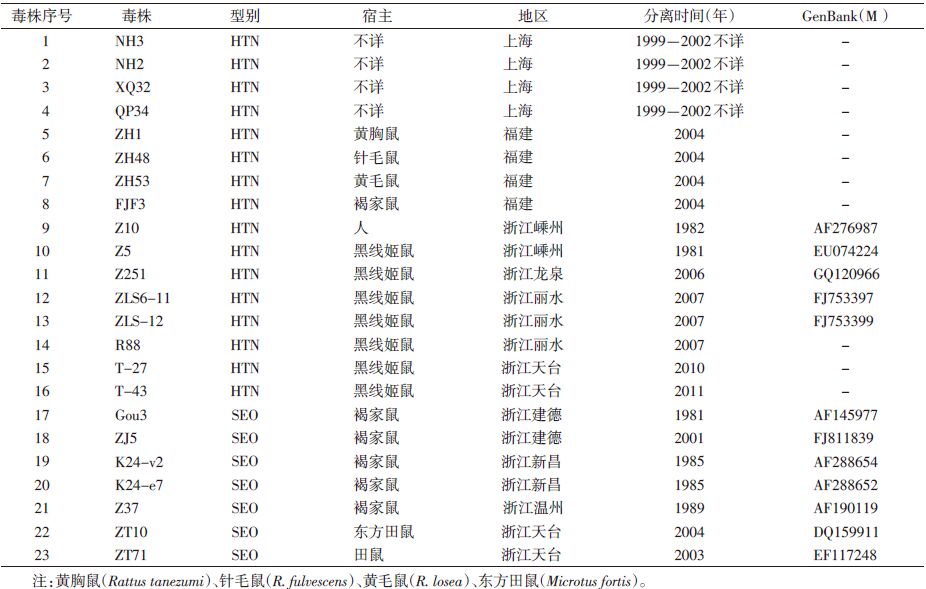
应用Mega 4.0和DNAStar软件包进行系统发生分析,以 NJ构建系统发生树,确定病毒的型别和基因差异程度。分析采用1000个多序列组。
2 结 果 2.1 汉坦病毒部分M片段核苷酸同源性上海HTN毒株NH3、QP34、XQ32、NH2之间的核苷酸同源性为87.3%~94.7%,而与HTNV国际标准株76-118之间的同源性为89.7%~99.7%;福建HTN毒株ZH1、ZH53、ZH48之间同源性为82.7%~95.7%,与76-118株同源性为85.7%~89.0%,而SEO毒株FJF3株与国际标准株80-39的同源性为93.3%;浙江株Ⅰ型毒株Z5、Z10、Z251、ZLS6-11、ZLS-12之间的同源性为86.6%~99.8%,与76-118同源性为84.2%~85.0%,SEO型毒株Gou3、ZJ5、K24-v2、ZT71、Z37、ZT10、K24-e7之间同源性为83.8%~99.7%,与80-39同源性为95.4%~84.2%,Gou3和ZJ5与80-39相差较大,分别为84.3%和84.2%,而他们自身较接近为91.2%。HTN的上海株与福建株同源性为84.3%~91.7%,与浙江株为 82.0%~86.3%,HTN的福建株与浙江株同源性为 80.7%~85.0%;SEO的福建株与浙江株同源性为84.0%~97.3%(图 1)。

|
| 图 1 东南沿海地区HV部分M片段核苷酸同源性 Figure 1 Comparison of HV nucleotide sequence homology of M in the South-east coastal area,China |
应用Mega 4.0和DNAStar软件包进行系统发生分析,以 NJ构建系统发生树,结果表明同型别病毒浙江省的毒株基本分布在临近分支,特别是浙江省同地区的毒株如丽水市的ZLS6-11、ZLS-12、R88和天台市的T-27、T-43,嵊州市的Z10、Z5,新昌市的K24-v2、K24-e7,建德市的Gou3、ZJ5和福建省的ZH1、ZH48均分布在同一分支上,表明上海市、福建和浙江省东南沿海地区汉坦病毒的基因差异和亲缘远近关系主要表现在地区性,显示出高度的地理聚集现象;同分离自浙江省而时间跨度相隔20多年的Z10和Z251、Gou3和ZJ5、Z37和ZT10分布在同一分支上,表明汉坦病毒的分离年代对病毒的变异不明显。基因差异在HTN发生群中,福建株ZH53独立分支;在SEO发生群中,浙江省建德市的Gou3和ZJ5独立进化支(图 2),表明福建和浙江省分别存在着HTN和SEO病毒基因变异较大、亲缘关系较远的特殊亚型病毒,而且在浙江省内部也存在着不同亚型的汉坦病毒。

|
| 图 1 东南沿海地区汉坦病毒部分M片段核苷酸系统发生树 Figure 1 Phylogenetic tree of HV based on M segment in the South-east coastal area,China |
本研究通过对浙江和福建省、上海市东南沿海地区汉坦病毒部分M片段的基因序列分析,并与汉坦病毒的HTNV和SEOV国际代表株76-118及80-39序列进行比较,表明东南沿海地区属HTN和SEO的混合疫区,不同型别的病毒对宿主具有相对选择性,HTNV和SEOV分别以黑线姬鼠和褐家鼠为主要宿主动物,但在福建省也存在着以黄毛鼠(Rattus losea)、黄胸鼠和针毛鼠为宿主的HTNV,在浙江省存在着以东方田鼠为宿主的SEOV[8],浙江省在2003-2004年从天台地区田鼠中分离到SEOV ZT10、ZT71株,而之前浙江省分离到的汉坦病毒主要来自于鼠亚科动物,未检测和分离到来自于田鼠亚科动物的汉坦病毒,表明浙江省内的汉坦病毒不断地进化突破了宿主之间间隔,产生宿主“溢出”现象。最近东南沿海的广东地区发现蝙蝠体内携带SEO的汉坦病毒,而且蝙蝠携带的与啮齿类动物携带的SEO汉坦病毒序列同源性达到99%以上[9],预示着可能因汉坦病毒由啮齿动物溢出到蝙蝠中,或者蝙蝠原本就是汉坦病毒的原始宿主。
在东南沿海地区的浙江、福建省、上海市HTNV和SEOV中,本省市分离的同型别毒株的基因序列同源性高于不同省市分离毒株,而本省市同地区分离株的核苷酸同源性更高,地区接近的分离株系统进化分支的分布也大部分更靠近,而同一地区不同年代的分离株基因差异不明显,表明病毒的基因差异和亲缘远近关系中主要表现在地区性,地理聚集现象明显,该现象在HTNV中表现尤为突出。
HTNV福建株ZH53与同型其他株及国际标准株76-118的核苷酸差异高于14.7%,在HTNV发生群中构成一独立分支;Gou3和ZJ5病毒分离自浙江省同一地区建德市,独立分布在同一个分支,但分离时间相距30年,互相之间的亲缘关系较近,在SEOV发生群中的独立分支以及与国内外其他SEOV基因的高度变异,这些现象显示在SEOV中病毒基因的变异空间比时间更起决定作用。浙江省建德地区存在着SEOV的特殊亚型病毒,提示建德地区独特的地理位置与这一结果的发现可能有关,建德市地处浙江西部,东接杭州市,西边黄山,中贯富春江、新安江,西北与千岛湖毗连。山陵与湖泊、江河的阻隔限制了宿主动物的迁移,进一步证明汉坦病毒的地理聚集现象[10]。
东南沿海的深圳市自2005年与本实验室合作分离研究汉坦病毒[11],研究发现深圳市汉坦病毒主要为褐家鼠体内携带的SEOV,M片段核苷酸同源性高达95.4%以上,属于同一亚型,同样说明汉坦病毒除具有宿主依赖性外,还具有地域的影响[12]。
本研究通过对东南沿海地区浙江、福建省和上海市不同年代不同宿主分离的汉坦病毒部分M基因序列的研究,表明东南沿海地区汉坦病毒基因差异和亲缘远近关系主要表现在地区性,而与病毒分离的年代与宿主关系并不大,表现出明显的地理聚集现象,该现象在HTNV中尤为明显,从而揭示东南沿海地区汉坦病毒基因的变异和进化规律,为HFRS的预防提供理论依据。
| [1] | 陈露菲,许军,刘忠伟,等. 黑龙江省首次从鼠肺中分离到家鼠型肾综合征出血热病毒[J]. 中国公共卫生,2001,17(4): 343. |
| [2] | Elliott RM,Schmaljohn CS,Collett MS. Bunyaviridae genome structure and gene expression[J]. Curr Top Microbiol Immunol, 1991,169(4):91-141. |
| [3] | 陈阳,李世清,何似. 福建省汉坦病毒基因分型及基因序列分析[J]. 中国人兽共患病学报,2004,20(10):855-857. |
| [4] | 李燕婷,周妍,周欣,等. 上海地区汉坦病毒基因分型和序列分析[J]. 上海预防医学杂志,2004,6(4):216-218. |
| [5] | 徐芳,朱函坪,姚苹苹,等. 浙江省2008-2011年啮齿动物中汉坦病毒的分离及鉴定[J]. 中国媒介生物学及控制杂志, 2013,24(4):285-288. |
| [6] | 姚苹苹,徐芳,朱函坪,等. 2007年浙江省鼠类感染汉坦病毒监测及病毒分离[J]. 中国媒介生物学及控制杂志,2009,20 (4):349-351. |
| [7] | 李婵,谢荣辉,朱函坪,等. 汉坦病毒Z5株M和S片段全基因序列测定及分析[J]. 中国人兽共患病学报,2010,26(3): 215-217. |
| [8] | 翁景清,谢荣辉,卢亦愚,等. 浙江东方田鼠分离汉坦病毒株S 基因的克隆和序列分析[J]. 中华微生物学和免疫学杂志, 2005,25(10):833-836. |
| [9] | 李敏,李林妙,张礼标,等. 广东省蝙蝠体内汉城型汉坦病毒的检测初报[J]. 广东农业科学,2014,41(8):177-180. |
| [10] | 姚苹苹,朱函坪,邓小昭,等. 浙江省汉坦病毒基因分子进化分析[J]. 病毒学报,2010,26(6):465-470. |
| [11] | 阳帆,古丽巴哈尔,刘建军,等. 深圳市2005年鼠间汉坦病毒感染的鼠情监测及病毒株SZ2083的分离鉴定[J]. 中华流行病学杂志,2006,27(11):981-984. |
| [12] | 刘建军,阳帆,何建凡,等. 深圳市鼠类感染汉坦病毒的分子特征[J]. 中华预防医学杂志,2008,42(5):324-328. |