 2015, Vol. 26
2015, Vol. 26扩展功能
文章信息
- 梁莹, 汪立茂, 骆孝志, 赵宏群, 蔡虹, 李伟
- LIANG Ying, WANG Li-mao, LUO Xiao-zhi, ZHAO Hong-qun, CAI Hong, LI Wei
- 四川省巴塘县首次分离鼠疫菌的遗传学特征
- Genetic characteristics of the first Yersinia pestis strain isolated from Batang county of Sichuan, China
- 中国媒介生物学及控制杂志, 2015, 26(3): 258-261
- Chin J Vector Biol & Control, 2015, 26(3): 258-261
- 10.11853/j.issn.1003.4692.2015.03.010
-
文章历史
- 收稿日期:2015-01-04




2. 四川省疾病预防控制中心, 成都610041;
3. 日喀则市疾病预防控制中心, 西藏日喀则857000
2 Sichuan Center for Disease Control and Prevention, Chengdu 610041, Sichuan Province, China;
3 Rikaze Center for Disease Control and Prevention, Rikaze 857000, Tibet Autonomous Region, China
鼠疫耶尔森菌(鼠疫菌)以啮齿动物为主要宿主,通过媒介跳蚤的叮咬进行传播,可引起自然疫源地内动物鼠疫的暴发流行,并可感染人类。鼠疫在我国的流行历史可追溯到公元6世纪的第一次世界鼠疫大流行时期,截止到2013年底,鼠疫自然疫源地分布于我国19个省(自治区)的301个县(市、旗),总面积达152万km2[1, 2]。其中,四川省已证实有2种类型的鼠疫自然疫源地,分别是青藏高原青海田鼠(Microtus fuscus)型疫源地和青藏高原喜马拉雅旱獭(Marmota himalayana)型疫源地,主要分布在甘孜州的石渠和德格县[3]。随着鼠疫监测工作的进一步开展,2011年又从甘孜州巴塘县境内的1只自毙旱獭体内分离到1株鼠疫菌,为进行疫源检索,协助判断这一新发现的鼠疫疫源地类型,本研究对分离自巴塘县的首株鼠疫菌进行了生化表型和基因型别的分析鉴定。 1 材料与方法 1.1 菌株来源
2011年在四川省甘孜州巴塘县境内发现1只自毙旱獭,从其体内分离到1株鼠疫菌,菌株随后由四川省疾病预防控制中心转运至本实验室保存,编号为川257。 1.2 生化实验
包括脱氮反应和糖醇酵解能力测定(甘油、鼠李糖、麦芽糖、阿拉伯糖),采用常规试管法。 1.2.1 培养基的制备
参照文献[4]中的配制方法,将每种培养基分装成小试管,高压灭菌,并经无菌实验后冷藏备用。 1.2.2 接种与培养
被试鼠疫菌株川257在赫氏消化液培养基上,28 ℃培养24 h,然后用一次性接种针挑取适量细菌培养物分别接种在以上5种鉴定用培养基中,37 ℃恒温孵箱中静置培养。糖醇发酵管需培养14 d,每天记录观察结果;脱氮反应管于接种2 d后,滴加第一液(0.8%的氨基苯磺酸醋酸溶液)和第二液(0.5%的α-萘胺醋酸溶液),根据颜色变化判定结果。实验同时设立空白对照。 1.3 基因型别鉴定
采用多位点可变数目串联重复序列分析(MLVA)的分子分型方法。 1.3.1 可变数目串联重复序列(VNTR)位点的选择
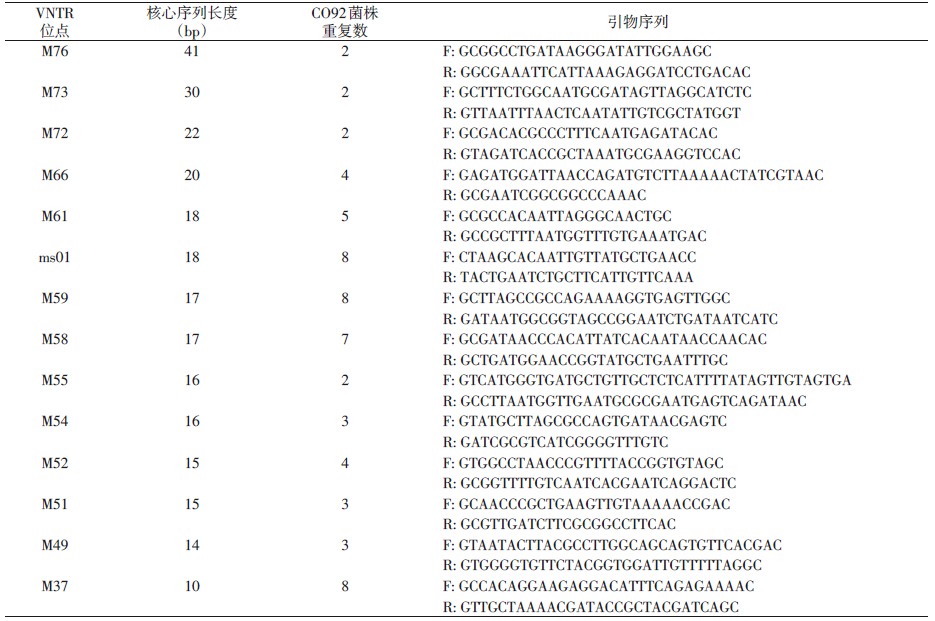
为便于菌株间亲缘关系的比较分析,选取文献[5]中报道的全部14个VNTR位点进行检测(表 1)。 1.3.2 引物
采用文献[5]中的引物序列(表 1),引物由生工生物工程(上海)有限公司合成。
 |
配制25 μl的反应体系,包括2×TransTaq-T PCR SuperMix(北京全式金生物技术有限公司)12.5 μl,上、下游引物各1 μl,2 ng/μl的DNA模板1 μl,以及超纯水9.5 μl。PCR扩增条件为94 ℃预变性5 min;94 ℃变性40 s,60 ℃退火40 s,72 ℃延伸50 s,35个循环;72 ℃再延伸10 min。 1.3.4 结果判定
2%琼脂糖凝胶电泳检测扩增产物的大小和产量,然后将PCR产物送深圳华大基因科技服务有限公司北京分部进行测序。将每个PCR产物的序列分别与参考菌株CO92的相应序列进行比对后确定菌株川257基因组中所有待测VNTR位点的重复数。 1.3.5 数据分析
选取55株分离自青藏高原喜马拉雅旱獭鼠疫自然疫源地的代表性鼠疫菌株,应用BioNumerics v5.10软件进行聚类分析,了解川257与这些菌株间的亲缘关系。 2 结 果 2.1 生化特性
菌株川257的主要生化表型为:甘油(+),脱氮(+),鼠李糖(-),麦芽糖(+),阿拉伯糖(+),根据鼠疫菌生物型和生态型的分型标准[6, 7],川257菌株为鼠疫菌的古典变种,其生态型为青藏高原型。 2.2 基因型别
琼脂糖凝胶电泳结果显示,用14对MLVA引物扩增菌株川257的基因组DNA均能得到单一、明亮的扩增产物条带(图 1),产物序列经比对后确定菌株川257在各个VNTR位点的重复数分别为M76(2)- M72(2)- M66(4)- M61(4)- ms01(7)- M59(7)- M58(7)- M55(2)- M54(4)- M51(3)- M49(3)- M37(7)。其余55株鼠疫菌的MLVA数据来自本实验室的鼠疫菌基因组多态性数据库。聚类分析结果表明,全部56株鼠疫菌可被分为3大群15个MLVA基因型(图 2),其中A群为青藏高原型和祁连山型鼠疫菌,群下包括0001~0010共10个MLVA基因型,来自青海、甘肃省和西藏自治区;B群全部为冈底斯山型鼠疫菌,群下包括0011~0014共4个MLVA基因型,菌株分离自横断山脉以南的藏南地区;C群全部为昆仑山A型鼠疫菌,群下只有1个MLVA基因型,即0015,菌株均来自新疆和田县。四川省巴塘菌株川257位于A群,MLVA基因型为0002,与其他3株分别分离自青海省贵德、尖扎、西藏自治区昌都的青藏高原型鼠疫菌的MLVA基因型完全相同。基因型0001和0003则全部为四川省德格的鼠疫菌分离株,与巴塘的川257等0002型菌株共同构成一个小亚群。

|
| 图 1 鼠疫菌株川257基因组中14个VNTR位点PCR扩增产物的琼脂糖凝胶电泳 Figure 1 Image of agarose gel electrophoresis of PCR amplification of 14 VNTR locus in the Y. pestis strain Chuan 257 |

|
| 图 2 基于14个VNTR位点多态性的鼠疫菌系统聚类图 Figure 2 Clustering dendrogram of Y. pestis strains based on 14 VNTR locus generated by Bionumerics v5.10 |
巴塘县位于四川省西部青藏高原东南缘,与2007年被证实为鼠疫自然疫源地的西藏芒康县分列金沙江中游的东、西两岸,其地形地貌、植被、气候、动物和媒介种群结构等地理生态环境均与芒康县十分相似。2010年巴塘县被列为四川省省级鼠疫监测点,随后在2011年的监测工作中即检出指示动物牧犬的鼠疫F1抗体阳性血清10份,并首次从喜马拉雅旱獭体内分离到鼠疫菌,从而确定巴塘县正在发生动物间鼠疫流行[8]。流行菌株的遗传学特征对疫源地性质判定、疫情溯源及合理配置人力物力进行鼠疫监测具有十分重要的意义。鼠疫菌在长期的传播扩散过程中,基因组不断发生变异,从而适应各种宿主、媒介、地理环境等不同的生态位类型。在我国目前已知的12种类型的鼠疫自然疫源地中,鼠疫菌自然分离株的遗传学特征各不相同,菌株致病性和毒力也强弱不等。
本研究利用MLVA分子分型方法,确定了在四川省巴塘县分离自喜马拉雅旱獭的首株鼠疫菌川257菌株基因组中14个VNTR位点的重复数特征,并据此与其他55株分离自青藏高原喜马拉雅旱獭鼠疫自然疫源地的代表性菌株进行了聚类分析,结果表明这些菌株的MLVA基因型与它们的生态型均能很好的相吻合,即同一个生态型的菌株,它们的MLVA位点特征也相同或十分相似,在聚类图上表现为聚集成群,如B群全部为冈底斯山生态型的菌株,C群全部为分离自新疆和田的昆仑山A型菌株,A群中虽然包括青藏高原型和祁连山型两种生态型的鼠疫菌株,但全部4株祁连山型菌株却自成一个小群。巴塘菌株川257与青海省贵德、尖扎和西藏自治区昌都的3株青藏高原型鼠疫菌的MLVA基因型完全相同,其生化表型特征也与青藏高原型菌株相一致,因此,可以确定巴塘菌株川257的生态型为青藏高原型。
早在20世纪90年代,巴塘县就发生过大量旱獭不明原因死亡,继而有人群被感染并死亡的事件,该县的地理环境特征与已知的藏北鼠疫疫源地极其相似,鼠疫菌的主要宿主和媒介也相同,而且两地之间没有高山峡谷等天然的地理屏障,结合现在的流行病学调查资料和菌株的遗传学特征鉴定结果,我们认为巴塘疫源地应该是青藏高原喜马拉雅旱獭鼠疫自然疫源地的扩展和延续。鼠疫菌很可能由青海省传入西藏横断山脉北部,再继续播散至四川省甘孜州的巴塘和德格等地。青藏高原生态型鼠疫菌的致病性是最强的,动物间鼠疫极易波及人间,为严防人间鼠疫发生,加强巴塘、德格及其毗邻各县的鼠疫监测和鼠防宣传教育工作刻不容缓。
| [1] | 卫生部卫生应急办公室,中国疾病预防控制中心. 鼠疫防控应急手册[M]. 北京:北京大学医学出版社,2009:1-2. |
| [2] | 张贵军,邵奎东,段天一,等. 全国2013年鼠疫监测结果[J]. 中国地方病防治杂志,2014,29增刊:9-17. |
| [3] | 汪立茂,祝小平,陈开华,等. 四川省鼠疫防治30年回顾[J]. 预防医学情报杂志,2012,28(1):1-9. |
| [4] | 夏连续,海荣. 鼠疫实验室技术手册[M]. 北京:北京出版社, 2013:24-26. |
| [5] | Zhang XA, Hai R, Wei JC, et al. MLVA distribution characteristics of Yersinia pestis in China and the correlation analysis[J]. BMC Microbiol,2009,9:205. DOI:10.1186/1471-2180-9-205. |
| [6] | Devignat R. Varieties of Pasteurella pestis:new hypothesis[J]. Bull World Health Organ,1951,4(2):247-263. |
| [7] | 纪树立,张海崚,刘云鹏,等. 我国鼠疫菌分型及其生态学、流行病学意义[J]. 中国地方病学杂志,1987,6(5):257-263. |
| [8] | 汪立茂,祁腾,杨军,等. 巴塘县为四川省又一旱獭鼠疫疫源县[J]. 预防医学情报杂志,2012,28(11):865-867. |